
Прокариотам принадлежит значительно большая роль в эволюции жизни, чем считалось ранееДо 1977 г. считалось, что ранние формы живых организмов и их современные потомки-прокариоты представляют собой непрерывную монофилетическую линию, в которой все формы сходны в отношении основ генетической и клеточной организации, а также механизмов экспрессии генов. Гипотеза эндосимбиоза. возрожденная в конце 1960-х гг. Л. Маргулис (Margulis) на основе более ранней гипотезы рубежа 19 и 20 вв., постулировала, что эукариотическая клетка — это химерная структура, возникшая при слиянии представителей различных эволюционных линий прокариот. Анализ последовательностей семантофорных молекул — вначале белков и затем рибосомных РНК — дал результаты, которые рассматриваются в пользу предположения Маргулис о происхождении органелл. В настоящее время считается, что митохондрии произошли из аэробных представителей Proteobacteria, хлоропласты — из цианобактерий.
Рис. 4. Ранняя концепция происхождения эукариот от прокариотического предка.
В настоящее время анаэробные бактерии-бродильщики рассматриваются как предки всех живых организмов в обеих предложенных концепциях эволюции прокариот — гипотезе конверсии,предложенной П. Броде (Brode), и гипотезе сегрегации,высказанной Маргулис. Согласно гипотезе сегрегации, эволюция путей транспорта электронов у анаэробно дышащих организмов предшествовала эволюции циклического транспорта электронов у анаэробных фототрофных и аэробно дышащих организмов. По этой гипотезе, каждое из данных свойств возникло лишь однажды в ходе эволюции; таким образом, группы всех современных бактерий-бродильщиков, анаэробно дышащих и фототрофных, хемоавтотрофных и аэробных типов организмов были помещены в раздельные эволюционные линии. Согласно гипотезе конверсии, фототрофные организмы возникли непосредственно из более древних организмов, осуществлявших брожение. В результате серии событий возникло несколько эволюционных линий, часть представителей которых вымерла, но часть существует до сих пор. Вследствие изменения со временем уровня кислорода в среде, от ранних фототрофных организмов взяли начало эволюционные линии, в которых развились различные типы метаболизма (например, органотрофия и литотрофия). Следовательно, анаэробы, фототрофы, анаэробно дышащие и аэробные формы развивались независимо одни от других несколько раз в ходе эволюции. Фундаментальные различия между этими двумя гипотезами касаются, таким образом, происхождения аноксигенного фотосинтеза и эволюции дыхания. Результаты филогенетического анализа в целом подтверждают гипотезу конверсии. Как отмечено выше, фототрофные бактерии встречаются в нескольких эволюционных линиях. Учитывая сложность фотосинтетического аппарата, разнообразие морфологических и функциональных типов и однообразие организации и принципа функционирования, есть основания предполагать, что фотосинтетический аппарат возник монофилетически, после чего его варианты сформировались независимо под действием отбора в определенных условиях обитания. Судя по положению рода Chloroflexus, наиболее далеко отстоящего на дереве, построенном по данным анализа 16S-рРНК, фотосинтетический аппарат возник на ранней стадии эволюции бактерий. Утрата фотосинтетического аппарата независимо у определенных представителей всех эволюционных линий, кроме двух (зеленых серных бактерий и цианобактерий), и изменения функции цепи циклического транспорта электронов в ответ на изменение условий питания привели к эволюции новых типов метаболизма. С гипотезой конверсии согласуются также сформировавшиеся в последнее время представления о том, что такие типы метаболизма, как брожение, анаэробное дыхание, хемоавтотрофия и аэробное дыхание, не эволюционировали монофилетически, но произошли от разных предковых форм в различных основных эволюционных линиях. Многие фототрофные бактерии, как установлено, близкородственны хемоавтотрофным и аэробным бактериям. Родство между Rhodopseudomonas palustris и окисляющим нитрит, строго аэробным видом Nitrobacter winogradskyi особенно ярко указывает на возможное возникновение дыхательной цепи из фотосинтетической цепи транспорта электронов. Следует, наконец, упомянуть, что гипотеза конверсии в целом согласуется с порядком ветвления деревьев, построенных на основе анализа последовательностей эволюционно консервативных макромолекул, но в деталях здесь возможны существенные расхождения.
7. Прокариоты составляют две основные эволюционные линии: домен Archaea и домен Bacteria В 1977 г. Вёзе и Фокс (Кох) путем анализа неполных последовательностей 16S- и 18S-рРНК установили, что прокариоты не представляют собой единую филогенетическую группу организмов. Данные анализа 16S-рРНК указали на две главные эволюционные линии, обозначенные как первичные царства. Молекулы 16S-рРНК у организмов каждой из этих главных эволюционных линий столь же не сходны между собой, как и с 18S-рРНК эукариот. «Типичные» бактерии, например Escherichia coli, были отнесены к царству бактерий, и несколько типов организмов, фенотипически отличающихся от бактерий, (табл.) — к царству архей. Последующее изучение рДНК простейших показало, что наиболее древние эукариотические организмы — это лямблии, микроспоридии и близкие к ним организмы. Как и прокариоты, эти простейшие обладают 16S-рРНК и лишены митохондрий. Раздельность данных трех основных эволюционных линий, установленную по результатам анализа рРНК, позже подтвердили результаты анализа генов некоторых белков, участвующих в процессах трансляции или получения энергии. Археи не составляют филогенетически единой группы и их следует распределить по трем эволюционным линиям: 1) галофилов, которые, возможно, относятся к (настоящим) бактериям (наличие «фотосинтетического» аппарата), 2) термофильных, ассимилирующих серу архей, называемых эоцитами и образующих недавно открытую, сильно разветвленную эволюционную линию, связанную с линией эукариот (действительно, существенное число «эоцитных» свойств имеется и у эукариот, но на их монофилетическое происхождение с археями указывают лишь немногие данные), 3) метаногенов, которые представляют собой истинные археи. Классификация с выделением трех царств, предложенная Вёзе, к настоящему времени заменена концепцией доменов,принятой в качестве предварительного варианта. Таксон домен был предложен как более высокий по отношению к царству, чтобы подчеркнуть значение подразделения мира живого на три части. Чтобы избежать впечатления, что суффикс «-bacteria» в названиях двух из трех царств отражает их особое эволюционное родство, были предложены термины Archaea и Bacteria взамен прежних Eubacteria и Archaebacteria и термин Eucarya вместо термина Eucaryota. В пользу самостоятельности Archaea как домена, столь же не родственного Eucarya и Bacteria, сколь неродственны два последних домена между собой, говорит также открытие того факта, что для Archaea характерен ряд особых эпигенетических свойств. Кроме того, специфические свойства и клеточные компоненты выявлены для определенных таксонов архей; это коферменты, участвующие в метаногенезе, пигменты пурпурных мембран, генерирующие энергию—бактериородопсин, галородопсин и другие сенсорные родопсины у галофилов, выживание в условиях экстремально высокой температуры, а также некоторые уникальные физиологические свойства (например, наличие модифицированного пути Энтнера-Дудорова). К уникальным для архей эпигенетическим характеристикам относятся следующие: • первичная структура рРНК и некоторых других семантофорных молекул, таких как АТР-синтаза и фактор элонгации EF-Tu; • вторичная структура рибосомных РНК; • клеточную стенку образует не характерный для бактерий пептидогликан; ею служит либо наружная мембрана, либо S-слой, состоящий из шести- или четырехугольно расположенных субъединиц белков или гликопротеинов (у всех Crenarchaeota, термофильных Euryarchaeota и Metanococcus), либо псевдомуреин (Metanobacterium) либо гетерополисахариды (галофилы); • устойчивость к пенициллину и D-циклосерину, обусловленная тем, что D-аминокислоты в клеточной стенке отсутствуют; • единственная ДНК-зависимая РНК-полимераза нечувствительна к антибиотикам, которые подавляют соответствующий фермент у бактерий; • природа мембранных липидов; полярные и неполярные липиды содержат разветвленные изопреноидные цепи, связанные с глицеролом простыми эфирными связями, а не сложноэфирными связями; • модификации тРНК.
|

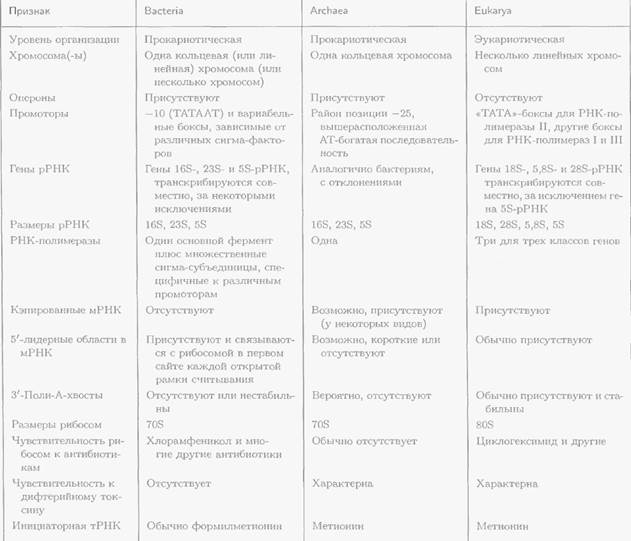 Таблица. Основные различия между эукариотами и двумя доменами прокариот (Bacteria и Archaea) на молекулярном уровне
Таблица. Основные различия между эукариотами и двумя доменами прокариот (Bacteria и Archaea) на молекулярном уровне


