В нижней части мозга расположен участок, называемый сосудодвигательным центром, который управляет кровообращением, а, следовательно, и кровяным давлением. Кровеносными сосудами, которые отвечают за контроль ситуации в системе кровообращения, являются артериолы, находящиеся между малыми артериями и капиллярами в кровеносной цепи. Сосудодвигательный центр получает информацию об уровне кровяного давления от нервов, чувствительных к давлению, которые располагаются в аорте и сонных артериях, а затем посылают сигналы к артериолам.
Практическая работа № 2
На тему:
Проводящая система сердца
Сделал: Ким Евгений Владимирович
Проверил: Архипова Татьяна Ивановна
Г.
Сокращение сердца.
Во время диастолы (Рисунок 4) предсердия (А) кровь течет от верхней и нижней полых вен в правое предсердие (1), а из четырех легочных вен - в левое предсердие (2). Поток увеличивается во время вдоха, когда отрицательное давление внутри грудной клетки способствует "присасыванию" крови в сердце, как воздуха в легкие. В норме это может проявляться дыхательной (синусовой) аритмией.
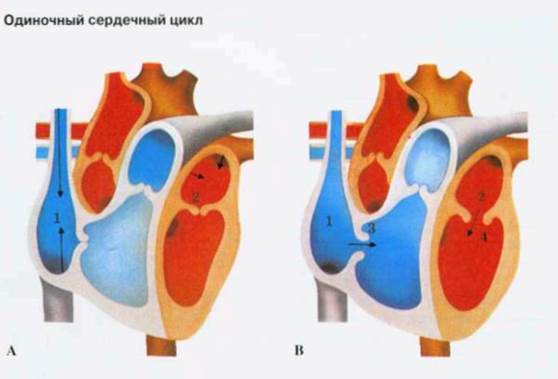
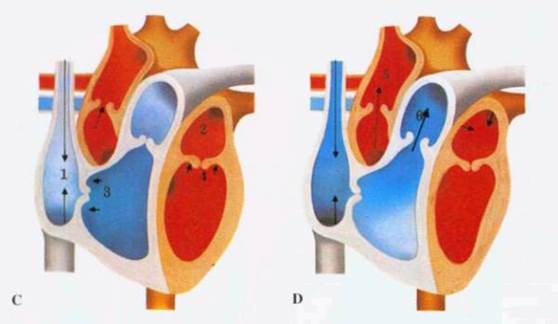
Рисунок 4.
Систола предсердий заканчивается (С), когда возбуждение достигает атриовентрикулярного узла и распространяется по ветвям пучка Гиса, вызывая систолу желудочков. Атриовентрикулярные клапаны (3, 4) быстро захлопываются, сухожильные нити и сосочковые мышцы желудочков препятствуют их заворачиванию (пролапсу) в предсердия. Венозная кровь заполняет предсердия (1, 2) во время их диастолы и систолы желудочков.
Когда систола желудочков заканчивается (В), давление в них падает, два атриовентрикулярных клапана - 3-створ-чатый (3) и митральный (4) - открываются, и кровь поступает из предсердий (1,2) в желудочки. Очередная волна возбуждения из синусного узла, распространяясь, вызывает систолу предсердий, во время которой через полностью открытые атриовентрикулярные отверстия в расслабленные желудочки нагнетается дополнительная порция крови.
Быстро возрастающее давление в желудочках (D) открывает аортальный клапан (5) и клапан легочного ствола (6); потоки крови устремляются в большой и малый круги кровообращения. Эластичность стенок артерий заставляет клапаны (5, 6) резко захлопываться в конце систолы желудочков.
Звуки, возникающие при резком захлопывании атриовентрикулярных и полулунных клапанов, выслушиваются через стенку грудной клетки как тоны сердца - "тук-тук".
Миокард предсердий и желудочков, разделенный фиброзными кольцами, синхронизируется в своей работе проводящей системой сердца, единой для всех его отделов (рис. 1.30).

Рис. 1.30. Схематическое изображение проводящей системы сердца: 1 — верхняя полая вена; 2 — синусно-предсердный узел; 3 — передний межузловой и межпредсердный тракт Бахмана; 4 — средний межузловой тракт Венкебаха; 5 — задний межузловой тракт Горела; 6 — предсердно-желудочковый узел; 7 — предсердно-желудочковый пучок; 8 — левая ножка предсердно-желудочкового пучка; 9 — правая ножка пучка Гиса; 10 — субэндокардиальная сеть волокон Пуркинье; 11 — нижняя полая вена; 12 — венечный синус; 13 — передняя ветвь левой ножки пучка Гиса; 14 — аорта; 15 — задний легочный ствол.
Структуры, генерирующие и передающие импульсы к предсердным и вентрикулярным кардиомиоцитам, регулирующие и координирующие сократительную функцию сердца, специализированы и сложны. Проводящая система сердца по своей гистоструктуре и цитологическим характеристикам существенно отличается от других отделов сердца. Анатомически проводящая система включает синусно-предсердный и предсердно-желудочковый узлы, межузловые и межпредсердные проводящие пути, предсердно-желудочковый пучок (пучок Гиса) специализированных мышечных клеток, отдающий левую и правую ножки, субэндокардиальную сеть волокон Пуркинье.
Синусно-предсердный узел
Синусно-предсердный узел расположен с латерильной стороны над основанием правого ушка у места впадения верхней полой вены в правое предсердие, от эндокарда которого его отделяет тонкая прослойка соединительной и мышечной ткани. Имеет форму уплощенного эллипса или полумесяца, горизонтально расположенного под эпикардом правого предсердия. Длина узла 10–15 мм, высота — до 5 мм, толщина — около 1,5 мм. Визуально узел слабо отличим от окружающего его миокарда, несмотря на капсулоподобное скопление соединительной ткани по периферии.
Ткань синусно-предсердного узла почти на 30% состоит из переплетающихся в различных направлениях пучков коллагеновых фибрилл различной толщины с небольшим количеством эластических волокон и клеток соединительной ткани. Тонкие мышечные волокна из специализированных клеток диаметром 3–4,5 мк расположены беспорядочно с неравномерными промежутками, выполненными интерстицием, микрососудами, нервными элементами, ориентированы по окружности сосуда, лишь вблизи центральной артерии, питающей узел. По периферии узел окружен значительным количеством фиброэластической ткани с обширной сетью капилляров, здесь же расположены нервные ганглии, единичные ганглиозные клетки и нервные волокна, в большом количестве проникающие в ткань узла.
Синусно-предсердный узел дает начало множественным путям, которые проводят импульсы, генерируемые специализироваными клетками. От него отходят латеральные пучки к правому ушку, нередко — горизонтальный пучок к левому ушку, задний горизонтальный пучок к левому предсердию и устьям легочных вен, пучки к верхней и нижней полым венам, медиальные пучки к межвенозному мышечному пучку миокарда. Данные мышечные пучки проводящей системы являются факультативными анатомическими образованиями, отсутствие того или иного из них может не оказывать заметного влияния на работу сердечной мышцы.
Межузловые пути проведения импульсов
Наиболее функционально значимыми являются нисходящие пути. Передний межузловой тракт, пучок Бахмана, берет начало от переднего края синусно-предсердного узла, проходит спереди и влево от верхней полой вены по направлению к левому предсердию, продолжаясь до уровня левого ушка. От пучка Бахмана ответвляется передний межузловой пучок, далее самостоятельно следующий в межпредсердной перегородке до предсердно-желудочкового узла. Средний меж узловой тракт, пучок Венкебаха, отходит от верхнего и заднего краев синусно- предсердного узла. Проходит единым пучком позади верхней полой вены, разделяясь затем на две неравные части, меньшая из которых следует до левого предсердия, а основная продолжается по межпредсердной перегородке до предсердно-желудочкового узла. Задний межузловой тракт, пучок Тореля, выходит из заднего края синусно-предсердного узла. Он рассматривается как основной путь межузлового проведения импульсов, его волокна следуют по пограничному гребешку, образуют основную долю волокон евстахиева гребня, следуя далее до предсердно-желудочкового узла по межпредсердной перегородке. Часть волокон перегородочной части всех трех трактов переплетается в непосредственной близости от предсердно-желудочкового узла, проникая в него на разных уровнях. Отдельные волокна межпредсердных и межузловых трактов по структуре сходны с волокнами Пуркинье желудочков, другие состоят из обычных предсердных кардиомиоцитов.
Предсердно-желудочковый узел
Предсердно-желудочковый узел обычно локализован под эндокардом правого предсердия на правом фиброзном треугольнике в нижней части межпредсердечной перегородки, над прикреплением септальной створки правого AV-клапана и несколько спереди от устья венечного синуса. Чаще всего овоидной, веретенообразной, дисковидной или треугольной формы, его размеры колеблятся в пределах от 6х4х05 до 11х6х1 мм.
В структуре предсердно-желудочкового узла, как и в рабочем миокарде, мышечный компонентпреобладает над соединительной тканью. В отличие от синусно-предсердного узла, он является мышечным образованием с менее развитым соединительнотканным остовом. Ткань узла как бы разграничена на две части крово снабжающей его артерией и пластинкой соединительной ткани, соединяющей стенку этого сосуда и фиброзное кольцо. От остальной ткани правого предсердия узел отделяет прослойка жировой клетчатки. Между предсердно-желудочковым узлом и устьем венечного синуса компактно размещены многочисленные парасимпатические ганглии. У мышечных волокон толщиной до 5 мкм продольное, косое и поперечное направление. Тесно переплетаясь, они образуют лабиринты, влияющие на электрофизиологические свойства ткани.
Пучок Гиса
От предсердно-желудочкового узла отходят верхний, задний и предсердно-желудочковый пучки Гиса, причем только последний выявляют в 100% наблюдений. Границей между пучком Гиса, отходящим от передней части предсердно-желудочкового узла, является его суженный участок, перфорирующий правый фиброзный треугольник в месте соединения с верхней перепончатой частью межжелудочковой перегородки. Длина пучка колеблется в пределах 8–20 мм при ширине 2–3 мм, толщине 1,5–2 мм и коррелирует с формой сердца.
По длиннику пучок Гиса слагается из двух частей: короткой интрафиброзной, проходящей сквозь ткань правого фиброзного треугольника, и более протяженной перегородочной, залегающей в межжелудочковой перегородке в виде серовато-бледного тяжа, который с возрастом приобретает желтоватый оттенок из-за накопления жировой ткани. На поперечных разрезах составляющие его мышечные волокна разделены соединительнотканными прослойками на группы, консолидированы в виде неправильного треугольника или фигуры овоидной формы. Предсердно-желудочковый пучок Гиса по всему периметру окружен плотной фиброзной тканью, размер его клеток возрастает по мере удаления от узла.
Под перепончатой частью, на уровне правого синуса аорты, пучок Гиса раздваивается на две ножки, как бы "седлая" гребень мышечного участка межжелудочковой перегородки. Более мощная правая ножка, сохраняющая вид пучка, проходит по правожелудочковой стороне межжелудочковой перегородки, отдавая ветви всем стенкам ПЖ. В большинстве случаев ее удается проследить до основания передней сосочковой мышцы, и лишь в отдельных наблюдениях она теряется уже на уровне середины межжелудочковой перегородки.
Топографически правая ножка пучка Гиса подразделяется на верхнюю, составляющую треть длины до основания перегородочных сосочковых мышц, среднюю — до перегородочно-краевой трабекулы, и нижнюю, расположенную в ней и в основании передней сосочковой мышцы. Верхняя часть этой ножки проходит субэндокардиально, следующая — интрамурально, а нижняя вновь возвращается под эндокард. Нижний участок ножки дает начало дистальным ветвям: передним, идущим к передней стенке желудочка, задним — к трабекулам задней стенки желудочка, и латеральным, следующим к правому краю сердца.
Левая ножка предсердно-желудочкового пучка появляется под эндокардом левой стороны межжелудочковой перегородки из-под задненижнего края перепончатой части перегородки между желудочками на уровне синусов аорты. В левой ножке различают стволовую и разветвленную части. Стволовая разделяется на переднюю ветвь, идущую к передней стенке ЛЖ и расположенной на ней сосочковой мышце, задняя — к его задней стенке и сосочковой мышце. При делении ножки на большее количество ветвей дополнительные ответвления следуют к верхушке сердца.
На периферии вторичные ветви левой ножки рассыпаются на более мелкие пучки, которые входят в трабекулы и образуют сетевидные связи между собой. Пучковые строения менее компактной левой ножки и обеих ее ветвей, направляющихся к передней и задней сосочковым мышцам, как и их граница с тканью рабочего миокарда, выражены значительно слабее, чем правой. Соединительнотканный и сосудистый компонент в них представлены хуже, чем в других участках проводящей системы. Клетки проводящей системы образуют под эндокардом сильно ветвящуюся сеть, элементы которой разграничиваются соединительнотканными прослойками, включающими сосудистые и нервные структуры.
Структура клеточных элементов
Строение клеток проводящей системы сердца определяется их функциональной специализацией. В ее неоднородном клеточном составе по морфофункциональным признакам выделяюттри типа специализированных кардиомиоцитов. Клетки I типа — П-клетки, типичные нодальные или ведущие пейсмейкерные — неправильной удлиненной формы. Эти небольшие миоциты диаметром 5–10 нм, со светлой саркоплазмой и довольно крупным центрально расположенным ядром отдают многочисленные цитоплазматические отростки, сужающиеся к концам и плотно переплетающиеся между собой. П-клетки образуют небольшие группы — кластеры, разграниченные элементами рыхлой соединительной ткани. Кластеры П-клеток окружены общей базальной мембраной толщиной 100 нм, глубоко проникающей в межклеточные щели. Их сарколемма образует многочисленные кавеолы, а вместо Т-системы — нерегулярно определяющиеся глубокие туннельные инвагинации диаметром 1–2 мкм, в которые проникает интерстиций и иногда — нервные элементы.
Контрактильный аппарат П-клеток представлен редкими, хаотично перекрещивающимися миофибриллами либо произвольно ориентированными свободно лежащими тонкими и толстыми протофибриллами и их пучками, нередко в комплексе с полирибосомами. Тонкие миофибриллы состоят из рыхло упакованных филаментов с небольшим количеством саркомеров, диски которых выражены нечетко, Z-линии неодинаковой толщины, иногда прерывисты, а электронно-оптически плотное вещество часто выходит за пределы миофибрилл. Объем, занимаемый миофибриллами в П-клетках, составляет не более 25% такового в вентрикулярных кардиомиоцитах. Редкие митохондрии неодинакового размера и формы с внутренней структурой, значительно упрощенной в сравнении с клетками рабочего миокарда, беспорядочно разбросаны в обильной светлой саркоплазме, окружающей относительно крупное ядро, которое расположено в центральной зоне. Гранулы гликогена немногочисленны.
Слабо развитый саркоплазматический ретикулум распределен преимущественно по периферии клетки, причем его терминальные цистерны иногда формируют типичные функциональные контакты с плазмолеммой. В цитоплазме содержатся свободные гранулы рибонуклеопротеидов, элементы гранулярного ретикулума, комплекса Гольджи, лизосомы. Стабильность формы этих довольно бедных органеллами клеток поддерживают многочисленные хаотично расположенные элементы цитоскелета — так называемые промежуточные филаменты диаметром около 10 нм, часто оканчивающиеся в плотном веществе десмосом.
Клетки II типа — переходные или латентные пейсмейкеры — неправильной удлиненной отростчатой формы. Они короче, но толще рабочих кардиомиоцитов предсердий, нередко содержат два ядра. Сарколемма переходных клеток часто образует глубокие инвагинации диаметром 0,12–0,16 мкм, выстланные гликокаликсом, как и в Т-тубулах. Эти клетки богаты органеллами и имеют меньше недифференцированной саркоплазмы, чем П- клетки, их миофибриллы ориентированы вдоль длинной оси, толще и состоят из большего количества саркомеров, в которых слабо выражены Н- и М-полоски. Митохондрии, расположенные между миофибриллами, по своей внутренней организации приближаются к таковым клеток рабочего миокарда, количество гликогена непостоянно.
Клетки III типа подобны клеткам Пуркинье — проводящие миоциты, на поперечных срезах выглядят объемнее других кардиомиоцитов. Их длина составляет 20–40 мкм, диаметр — 20– 50 мкм, образуемые ими волокна имеют большее поперечное сечение, чем в рабочем миокарде, но неодинаковую толщину.
Клетки Пуркинье отличают также обширная свободная от миофибрилл перинуклеарная зона, выполненная светлой вакуолизированной саркоплазмой, крупное округлое либо напоминающее прямоугольник ядро с умеренной концентрацией хроматина. Их контрактильный аппарат развит слабее, а система пластического обеспечения — лучше, чем в вентрикулярных кардиомиоцитах. Сарколемма образует многочисленные кавеолы, единичные, нерегулярно расположенные Т-тубулы и глубокие, достигающие аксиальной зоны клетки-туннели диаметром до 1 мкм, выстланные базальной мембраной.
Миофибриллы, расположенные в субсарколеммной зоне, иногда ветвятся и анастомозируют. Несмотря на нечеткую ориентировку по длиннику клетки, они, как правило, закреплены в обоих вставочных дисках. Упаковка миофиламентов в миофибриллах довольно рыхлая, гексагональное расположение толстых и тонких протофибрилл не всегда выдерживается, в саркомерах слабо выражены Н- полоска и мезофрагма, отмечается полиморфизм в структуре Z-линий.
В саркоплазме видны свободно взвешенные разрозненные и собранные в комплексы толстые и тонкие филаменты цитоскелета, связанные с полисомами, микротрубочки, лептофибриллы с периодом 140–170 нм, рибосомы и гранулыгликогена, нередко заполняющие всю свободную саркоплазму. Немногочисленные элементы саркоплазматического ретикулума располагаются вокруг миофибрилл и под сарколеммой, иногда образуют субсарколеммные цистерны. Митохондрии заметно меньше, чем в рабочих кардиомиоцитах, расположены как вдоль миофибрилл, так и перинуклеарно в виде небольших скоплений. Здесь же отмечаются профили гранулярного ретикулума, пластинчатого комплекса, лизосомы, окаймленные везикулы.
В целом, П-клетки проводящей системы, генерирующие импульсы, отличаются наиболее низким уровнем морфологической дифференцировки, который постепенно повышается по мере приближения к рабочим кардиомиоцитам желудочков, достигая здесь максимального значения. Объединение различных типов клеток в единую систему генерации и проведения импульса определяется необходимостью синхронизации этого процесса во всех отделах сердца.
Миоциты проводящей системы сердца имеют не только цитоморфологические, но иммуно- и гистохимические отличия от клеток рабочего миокарда. Все миоциты проводящей системы, за исключением П-клеток предсердно-синусного узла, богаче гликогеном, который присутствует в них не только в легко метаболизируемой β-форме, но и в виде более устойчивого комплекса с белками — десмогликогена, выполняющего пластические функции. Активность гликолитических ферментов и гликогенсинтетазы в проводящих кардиомиоцитах относительно выше, чем энзимов цикла Кребса и дыхательной цепи, тогда как в рабочих кардиомиоцитах это соотношение имеет обратный характер соответственно содержанию митохондрий. В результате миоциты предсердно-желудочкового узла, пучка Гиса и других отделов проводящей системы устойчивее к гипоксии, чем остальной миокард, несмотря на более высокую активность АТФазы. В ткани проводящей системы отмечается интенсивная реакция на холин эстеразу, отсутствующая в миокарде желудочков, и значительно большая активность лизосо мальных гидролаз.
Распределение миоцитов различных типов, характер и строение контактов клеток в различных отделах проводящей системы определяется их функциональной специализацией. В срединной зоне синусно-предсердного узла расположены наиболее рано активирующиеся П-клетки — пейсмейкеры, генерирующие импульс. Его периферию занимают переходные клетки II типа, П-клетки контактируют только с ними. Переходные клетки опосредуют прохождение импульса к миоцитам предсердий, замедляют его распространение. Контакты П-клеток немногочисленны, имеют упрощенное строение и весьма произвольную локализацию. В большинстве случаев представлены простым сближением плазмолеммы смежных клеток, фиксируемым единичными десмосомами. Цитологический состав предсердно-желудочкового узла более разнообразен. В нем присутствуют клетки, по структуре очень близкие к пейсмейкерным, краниодорсальную часть занимают миоциты II типа, а дистальные отделы состоят из быстрее проводящих импульс Пуркинье-подобных проводящих миоцитов III типа.
Некоторые исследователи выделяют в составе узла три зоны, отличающиеся по морфологическим и электрофизиологическим характеристикам: АN, переходную от предсердного миокарда к узловой ткани, состоящую в основном из переходных клеток, и NН-зону, пограничную с пучком Гиса, преимущественно формируемую полиморфными переходными Пуркинье-подобными клетками.
Контакты переходных миоцитов с типичными нодальными П-клетками имеют более простое строение, чем их соединения между собой, с предсердными рабочими миоцитами или клетками III типа. Межклеточные стыки образуют лишь непротяженные и бедные осмиофильным материалом промежуточные зоны, а десмосомы и миниатюрные нексусы отмечают довольно редко.
Межклеточные контакты миоцитов III типа между собой и с окружающими сократительными кардиомиоцитами организованы сложнее и по своей структуре ближе к характерным для рабочего миокарда. Вследствие более упорядоченного расположения миофибрилл они ориентированы поперек длинной оси клеток и замет- но реже образуются боковыми поверхностями их апикальных зон. Поперечно расположенные вставочные диски отличает большая протяженность хорошо выраженных промежуточных зон. Наличие протяженных нексусов при боковых контактах значительно повышает проводимость этих мышечных волокон и облегчает передачу импульсов на рабочий миокард. Вставочные диски между клетками Пуркинье иногда имеют косое расположение или V-образную форму. Подобная ориентация и слабая извитость промежуточных зон соответствуют более примитивному строению их вставочных дисков по сравнению с рабочими клетками.




