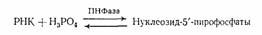Макромолекулярная структура ДНК.
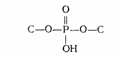
Длина витка спирали, который соответствует ее периоду идентичности, составляет 3,37 нм (33,7 А). На один виток спирали приходится 10 остатков оснований в одной цепи. Расстояние между плоскостями оснований равно, таким образом, примерно 0,34 нм (3,4 А). Плоскости остатков оснований перпендикулярны длинной оси спирали. Плоскости углеводных остатков несколько отклоняются от этой оси (первоначально Уотсон и.Крик предположили, что они параллельны ей). Из рисунка видно, что углеводофосфатный остов молекулы обращен наружу. Спираль закручена таким образом, что на ее поверхности можно выделить две различные по размерам бороздки (их часто называют также желобками) — большую, шириной примерно 2,2 нм (22 А), и малую —шириной около 1,2 нм (12А). Спираль — правовращающая. Полидезоксирибонуклеотидные цепи в ней антипараллельны: это означает, что если мы будем двигаться вдоль длинной оси спирали от одного ее конца к другому, то в одной цепи мы будем проходить фосфодиэфирные связи в направлении 3' - 5', а в другой — в направлении 5' - 3'. Иными словами, на каждом из концов линейной молекулы ДНК расположены 5'-конец одной и 3'-конец другой цепи. Регулярность спирали требует, чтобы против остатка пуринового основания в одной цепи находился остаток пиримидинового основания в другой цепи. Как уже подчеркивалось, это требование реализуется в виде принципа образования комплементарных пар оснований, т. е. остаткам аденина и гуанина в одной цепи соответствуют остатки тимина и цитозина в другой цепи (и наоборот). Таким образом, последовательность нуклеотидов в одной цепи молекулы ДНК предопределяет нуклеотидную последовательность другой цепи. Этот принцип является главным следствием модели Уотсона и Крика, поскольку он в удивительно простых химических терминах объясняет основное функциональное назначение ДНК — быть хранителем генетической информации. Заканчивая рассмотрение модели Уотсона и Крика, остается добавить, что соседние пары остатков оснований в ДНК, находящейся в В-форме, повернуты друг относительно друга на 36° (угол между прямыми, соединяющими атомы С1' в соседних комплементарных парах). 5. Природа межнуклеотидных связей Работы по определению способа соединения нуклеотидов в полимерных молекулах НК были успешно завершены в начале 50-х годов сразу после того, как была установлена структура нуклеотидов и изучены некоторые свойства их производных (главным образом эфиров). К этому же времени были разработаны методы выделения и очистки ДНК и РНК, так что исследование природы межмономерных связей проводилось с использованием чистых, хотя и сильно деградированных препаратов НК. Первые сведения о типе межмономерной, или, как ее принято называть, межнуклеотидной связи были получены с помощью потенциометрического титрования. Эти сведения свидетельствовали о наличии как в РНК, так и в ДНК только одной гпдроксильной группы у каждой фосфатной группы (рКа~1). На основании этого было сделано заключение, что НК содержит структурную единицу дизамещенной фосфорной кислоты. Естественно было предположить, что фосфатные остатки «сшивают» нуклеозиды за счет двух своих гидроксилов, а один остается свободным. Оставалось выяснить, какие части нуклеозидных фрагментов участвуют в образовании связи с фосфатными группами. Поскольку НК могут быть дезаминированы действием азотистой кислоты, очевидно, что аминогруппы пиримидиновых и пуриновых оснований не принимают участия в образовании межнуклеотидной связи. Помимо этого потенциометрическое титрование указывало, что и оксо(окси)-группы остатков гуанина и урацила, входящих в состав НК, свободны. На основании этих данных было сделано заключение о том, что межнуклеотидные связи образованы фосфатной группой и гидроксильными группами углеводных остатков (т. е. что они являются фосфодиэфирными), которые, следовательно, и являются ответственными за образование полимерной цепи (НК). Таким образом, то, что принято обычно называть межнуклеотидной связью, представляет собой по существу узел, включающий систему связей:
(где С — первичный или вторичный атомы углерода остатка углевода). При гидролизе ДНК и РНК в зависимости от условий реакции, образуются нуклеотиды с разным положением фосфатного остатка:
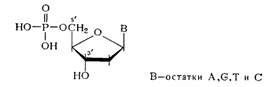
Если предположить, что в НК все межнуклеотидные связи идентичны, то, очевидно, что они могут включать помимо фосфатного остатка только З'-гидроксильную группу одного нуклеозидного звена и 5'-гидроксильную группу другого нуклеозидного звена (3'—У-связь). В случае же их неравноценности в полимерной цепи ДНК могли бы одновременно существовать три типа связей: 3'—5', 3'—3' и 5'—5'. Для РНК за счет участия 2/-гидpoкcилыIoй группы число типов связи должно было быть еще больше. Установить истинную природу межнуклеотидных связей в нативных ДНК и РНК удалось в результате направленного расщепления биополимеров с помощью химического и ферментативного гидролиза и последующего выделения и идентификации полученных при этом фрагментов. 5.1. Межнуклеотидная связь в ДНК Химический гидролиз ДНК как метод деградации полимера с целью установления природы межнуклеотидной связи оказался практически непригодным. ДНК не расщепляется при щелочных значениях рН, что хорошо согласуется с предположением о фосфодиэфирной природе межнуклеотидной связи (устойчивость диалкилфосфатов в щелочной среде обсуждалась в разделе). При обработке кислотой даже в мягких условиях ДНК расщепляется как по фосфодиэфирным, так и по N-гликозидным связям, образованным пуриновыми основаниями. Вследствие этого расщепление полимера протекает неоднозначно, но из продуктов кислотного гидролиза ДНК все же удалось выделить ди-фосфаты пиримидиновых дезоксинуклеозидов, которые оказались идентичными синтетическим 3',5'-дифосфатам дезоксицитидина и дезокситимидина:
Здесь же важно отметить, что наличие этих соединений в продуктах деградации ДНК указывает на участие обеих гидроксильных групп, по крайней мере пиримидиновых мономерных компонентов, в образовании межнуклеотидной связи. Более специфическим оказалось ферментативное расщепление ДНК. При обработке препаратов ДНК фосфодиэстеразой style='mso-spacerun:yes'> (ФДЭ) змеиного яда полимер практически полностью гидролизуется до дезоксннуклеозид-5'-фосфатов, структура которых была Остановлена сравнением с соответствующими нуклеотидами, полученными встречным синтезом.
Эти данные свидетельствуют об участии 5'-гидроксильных групп всех четырех дезоксинуклеозидов, входящих в состав ДНК, в образовании межнуклеотидной связи. Аналогично, но до 3'-фосфатов дезоксинуклеозидов расщепляется ДНК в присутствии ФДЭ, выделенной из Микрококков или из селезенки.
Из данных гидролиза ДНК фосфодиэстеразами различной специфичности становится очевидным, что связь нуклеозидных остатков в ДНК осуществляется фосфатной группой, которая одновременно этерифицирует гидроксильную группу у вторичного атома углерода (положение 3') одного нуклеозидного звена и гидроксильную группу у первичного атома углерода (положение 5') - другого нуклеотидного звена. Таким образом, было убедительно доказано, что в ДНК межнуклеотидная связь осуществляется за счет фосфатной группы, а также 3'- и 5'-гидроксильных групп нуклеозидных остатков [(а) и (б) — направления расщепления полинуклеотидной цепи ДНК фосфодиэстеразами соответственно змеиного яда и селезенки или микрококков]:
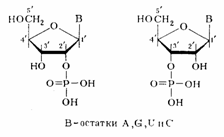
Предположение о возможности иного строения полимера с регулярно перемежающимися связями нуклеозидных остатков по типу 3'—3' и 5'—5' было отвергнуто, так как оно не удовлетворяло всем экспериментальным данным. Так, полимер такого типа не должен был бы полностью гидролизоваться (до мономеров) в присутствии ФДЭ змеиного яда, избирательно расщепляющей только алкиловые эфиры нуклеозид-5' –фосфатов. То же можно сказать о ФДЭ селезенки, селективно гидролизирующей алкиловые эфиры нуклеозид-3'-фосфатов. 5.2. Межнуклеотидная связь в РНК Более сложным оказался вопрос о природе межнуклеотидной связи в РНК. Уже на первых этапах изучения строения РНК был установлен факт чрезвычайной неустойчивости се при щелочном гидролизе. Основными продуктами щелочного гидролиза РНК являются рибонуклсозид-2'- и рибонуклеозид-З'-фосфаты, образующиеся практически в равных количествах.
Рибонуклеозид-5'-фосфаты при этом не образуются. Эти данные не укладывались в представления о фосфодиэфирной природе межнуклеотидной связи в РНК и требовали всестороннего изучения. Очень важную роль в таком исследовании, которое выполнили в начале 50-х гг. Тодд с сотрудниками, сыграли синтетические алкиловые эфиры рибонуклеотидов, которые были получены специально, чтобы промоделировать тот или иной тип фосфодиэфирной связи. Полученные школой Тодда данные о механизмах превращения алкиловых эфиров рибонуклеотидов в щелочной среде позволили предположить, что в РНК, так же как и в ДНК, межнуклеотидная связь осуществляется фосфатной группой и 3'- и 5'-гидроксильными группами углеводных остатков. Подобная связь в РНК должна очень легко расщепляться в щелочной среде, так как соседняя 2'-гидроксильная группа должна катализировать этот процесс при рН>10, когда начинается ионизация гидроксильных групп рибозы. Очень важно подчеркнуть, что промежуточными соединениями при щелочном расщеплении должны быть все четыре рибонуклеозид-2',З'-циклофосфата, а конечными — образующиеся при их гидролизе рибонуклеозид-3'-фосфаты и рибонуклеозид-2'-фосфаты (четыре пары изомеров). Данные щелочного гидролиза ограничили количество возможных для РНК типов межнуклеотидных связей, но не прояснили вопроса о том, как построен этот полимер. Более точные сведения о типе межнуклеотидной связи в РНК, как и в случае ДНК, были получены с помощью ферментативного гидролиза. Гидролиз РНК с использованием ФДЭ змеиного яда, протекающий до рибонуклеозид-5'-фосфатов, подтвердил уже прямым путем предположение об участии 5'-гидроксильных групп в образовании фосфодиэфирной связи между мономерными звеньями. Позднее это было окончательно установлено в результате открытия фосфоролиза РНК в присутствии фермента полинуклеотидфосфорилазы (ПНФаза), приводящего к образованию рибонуклеозид-5'-пирофосфатов:
Таким образом, оставалось выяснить природу второй гидроксильной группы, участвующей в образовании межнуклеотидной связи. Частично решить эту задачу помог еще один фермент, который использовался для направленного расщепления РНК, — пиримидиловая рибонуклеаза (РНаза). Ранее было показано, что этот фермент расщепляет только алкиловые эфиры пиримидиновых рибонуклеозид-3'-фосфатов до рибонук-леозид-3'-фосфатов (через промежуточный рибонуклеозид-2',З'-циклофосфат). Оказалось, что аналогичным образом этот фермент действует и на РНК. В экспериментах с любыми образцами очищенной РНК было обнаружено, что количество фосфорной кислоты, которая образуется при обработке полимера последовательно пиримидиловой РНазой и фосфомоноэстеразой (ФМЭ), а также количество иодной кислоты, затрачиваемой на последующее окисление, эквивалентно количеству остатков пиримидинов в данном образце РНК. Это говорило в пользу того, что по крайней мере пиримидиновые нуклеотиды в РНК связаны с соседними нуклеотидами только посредством 3'—5'-межнук-леотидной связи. Этот вывод подтверждают данные щелочной обработки ферментативных гидролизатов РНК, полученных после действия на нее РНазы: в щелочной среде миграция фосфатного остатка в рибонуклеозид-З'- и -2'-фосфатах невозможна, и наличие в соответствующих гидролизатах только пиримидиновых рибонуклеозид-З'-фосфатов делает очевидным 3'—5'-тип межнуклеотидной связи для пиримидиновых нуклеотидов. 6. Значение нуклеиновых кислот Значение нуклеиновых кислот очень велико. Особенности их химического строения обеспечивают возможность ранения, переноса в цитоплазму и передачи по наследству дочерним клеткам информации о структуре белковых молекул, которые синтезируются в каждой клетке. Белки обусловливают большинство свойств и признаков клеток. Понятно поэтому, что стабильность структуры нуклеиновых кислот - важнейшее условие нормальной жизнедеятельности клеток и организма в целом. Любые изменения строения нуклеиновых кислот влекут за собой изменения структуры клеток или активности физиологических процессов в них, влияя таким образом на жизнеспособность. Существует два типа нуклеиновых кислот: ДНК и РНК. ДНК (дезоксирибонуклеиновая кислота) - биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырех азотистых оснований: аденин(А) или тимин (Т), цитозин (Ц) или гуанин (Г); пятиатомный сахар пентозу - дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов. В каждой цепи нуклеотиды соединяются путем образования ковалентных связей между дезоксирибозой одного и остатком фосфорной кислоты последующего нуклеотида. Объединяются две цепи в одну молекулу при помощи водородных связей, возникающих между азотистыми основаниями, входящими в состав нуклеотидов, образующих разные цепи. Количество таких связей между разными азотистыми основаниями неодинаково и вследствие этого они могут соединяться только попарно: азотистое основание А одной цепи полинуклеотидов всегда связано двумя водородными связями с Т другой цепи, а Г - тремя водородными связями азотистым основанием Ц противоположной полинуклеотидной цепочки. Такая способность к избирательному соединению нуклеотидов называется комплиментарностью. Комплиментарное взаимодействие нуклеотидов приводит к образованию пар нуклеотидов. В полинуклеотидной цепочке соседние нуклеотиды связаны между собой через сахар и остаток фосфорной кислоты. РНК (рибонуклеиновая кислота), так же как ДНК, представляет собой полимер мономерами которого служат нуклеотиды. Азотистые основания те же самые, что входят в состав ДНК (аденин, гуанин, цетозин); четвертое - урацил - присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК содержат вместо дизоксирибозы другую пентозу - рибозу. В цепочке РНК нуклеотиды соединяются путем образования ковалентных связей между рибозой одного нуклеотида и остатком фосфорной кислоты другого. Известны двух- и одноцепочные молекулы рибонуклеиновой кислоты. Двухцепочные РНК служат для хранения и воспроизведения наследственной информации у некоторых вирусов, т.е. выполняют у них функции хромосом. Одноцепочные РНК осуществляют перенос информации о последовательности аминокислот в белках от хромосомы к месту их синтеза и участвуют в процессах синтеза. Существует несколько видов одноцепочных РНК. Их названия обусловлены выполняемой функцией или местом нахождения в клетке. Основную часть РНК цитоплазмы (80-90%) составляет рибосомальная РНК (рРНК). Она содержится в органоидах клетки, осуществляющих синтез белков, - рибосомах. Размеры молекул рРНК относительно невелики, они содержат от 3 до 5 тысяч нуклеотидов. Другой вид РНК - информационные (иРНК), переносящие от хромосом к рибосомам информацию о последовательности аминокислот в белках, которые должны синтезироваться. Транспортные РНК (рРНК) включают 76-85 нуклеотидов и выполняют несколько функций. Они доставляют аминокислоты к месту синтеза белка, «узнают» (по принципу комплиментарности) участок иРНК, соответствующий переносимой аминокислоте, осуществляет аминокислоты на рибосоме. Материалы с сайта http://km.ru
|