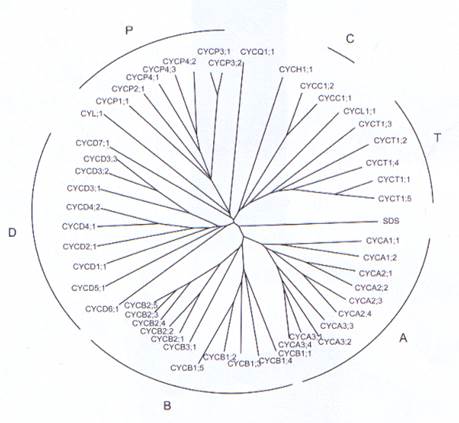
Растительные циклины2.1. Вступление Циклины являются регуляторные партнеры циклинзависимых киназ (CDK) – циклиновых гетеродимерных протеинкиназных комплексов. Открытие циклинов в яйцах морского ежа Тимом Хантом (Tim Hunt) и его коллегами в 1983 году дало ключевую подсказку для понимания биохимических механизмов, лежащих в основе осциллятора, что управляет клеточным циклом и раскрыло базовые клеточные механизмы, свойственные всем эукариотам. В оригинальной работе по открытию циклинов были описаны только два циклина, имеющие отношение к S-фазе (А-тип) и M-фазе (В-тип), которые, как было показано позднее, связываются с одной субъединицой киназы, оказавшейся Cdk 1. Сегодня известна большая группа животных, дрожжевых и растительных циклинов, формирующих связи с большим количеством разных СDK, так же циклиновые комплексы признаны ключевым видом регуляции серин-треониновых протеинкиназ. Связывание циклинов – мощная стратегия, поскольку СDK инактивированы без циклинового комплекса, так что регулируемый синтез, разрушение или локализация циклиновых субъединиц обеспечивает множественные уровни контроля для циклиновой активации субъединиц CDK так же хорошо, как контроль возможных комплексов различных циклинов и CDK. Этот гегуляторный модуль очень комбинаторный, но неясно точно его использование в контроле клеточного цикла, с большим количеством других примеров СDK-циклиновой регуляции, включая транскрипцию и ответа на наличие фосфата. 2.1.1 Циклины и осциллятор клеточного цикла Основной принцип осциллятора клеточного цикла такой, что регулируемый синтез и разрушение циклинов обеспечивает колебания CDK-активности, что управляет двумя ключевыми переходами в клеточном цикле – переход из G1 в S фазу и из G2 в М фазу. Теоретически, можно утверждать, что только одного циклина может быть достаточно для работающего осциллятора клеточного цикла и, в самом деле, Фишер и Нюрс (Fisher and Nurse) доказали на дрожжах, что один циклин способен содействовать ДНК-синтезу и митозу. Осциляторная модель одного циклина, впрочем, была опровергнута из-за чрезмерной упрощенности, так как впоследствии было открыто много различных циклинов. Более того, отчасти иные принципы, лежащие в основе циклиновой регуляции двух основных контрольных точек. G1-S циклины часто реагируют на внешние сигналы, такие как гормоны, или внутренние сигналы, такие как темпы роста или размер клетки, и являются обычно нестабильными белками, быстро оборачивающимися в убиквитинилирование-протеосомном пути. Комбинация регуляции экспрессии такими сигналами ведет к их быстрому исчезновению на удаление этих стимулов. Такое поведение типично для D-типа циклинов, которые потом передаются потом в клеточный цикл в удобном состоянии для деления и как таковые «установливаются на болтах» к ядру осциллятора, обеспечивая возможность для клеточного, окружающего(экологиечского) и эволюционного наблюдения за началом клеточного цикла. С другой стороны, S-фаза (циклин А) и митотические циклины (циклин В) демонстрируют регулируемый синтез и разрушение на зависимый от клеточного цикла манер и их изобилие скорее зависит от стадии клеточного цикла, чем от внешних сигналов. У животных было идентифицировано по крайней мере 13 классов циклинов (называемых циклинами от A до L и T), в растениях более, чем 100 циклинов были выделены из различных растений. Обширный геномный анализ Арабидопсиса дал 50 предполагаемых циклинов, которые можно сгруппировать в 9 классов, из которых 5 классов не были найдены у животных, хотя точно доказано, что для клеточного цикла существует роль только для циклинов А, В, D и Н. Читатель должен помнить, что циклины определяются, как белки, содержащие консервативную часть, называемую циклиновым боксом (cyclin box), регион, несущий СDK-связывающий сайт. Однако, это не значит, что каждый белок с этим доменом имеет функцию в ядре машинерии клеточного цикла. Изобилие циклинов отображает тот факт, что развитие комплекса многоклеточных организмов требует пространственного и временного контроля клеточного деления. И растения, и животные нуждаются в модуляции клеточного деления для того, что бы следовать своей схеме развития и регулировать свой исходный рост для адаптации к меняющимся условиям внешней среды. Растения, в особенности, нуждаются в приспособлении их неопределенной модели роста на гибкий манер, так как они не способны к движению в более оптимальное место. Большая часть современных знаний о биохимических механизмах регуляции клеточного цикла получена от исследований животных и дрожжей и мы только начинаем раскрывать аналоги и специфичные для растений регуляторные механизмы в растительном клеточном цикле. Эта часть расскажет о прогрессе в изучении циклиновых белков. 2.2 Семья растительных циклинов Гомология позволяет терминологии растительных циклинов быть стандартизированнй международным соглашением, как описано, и все растительные циклины были названы соответственно их предложениям. В названиях используются, когда необходимо, две буквы, описывающие вид, так первая буква рода и вида (напр. At для Arabidopsis thaliana), следующие за тремя буквами СYC и далее буква, обозначающая основную группу циклинов, которые определены по гомологии с другими высшими эукариотами. Далее следует номер подгруппы, напр. СYCA3 (не сохраняется вне растений), и видовой номер гена, где представлены множественные гены в одном виде растения (напр. СYCA3;3). Этот последний номер гена не сохраняется между видами. 2.2.1 Филогенетические связи между циклинами животных и растений Растительные циклины частично гомологичны с А-, В-, С-, Н- и L-типами классов циклинов млекопитающих, хотя подклассы в пределах этих классов не схожи между растениями и животными. Гомологии между Арабидопсисными и человеческими Е-, F-, G-, I- и UNG2-циклинами не существует. В Arabidopsis, 50 гомологов циклина были разделены в 9 классов с 23 подклассами и двумя дополнительными циклиновыми белками, не относящимися ни к одному классу (Рис. 2.1.) Эти «сиротливые» циклины - CYCJ18, теперь называемый как СYCQ1;1, и циклин-подобный белок CYL;1.
Рис. 2.1 Филогенетическое дерево анализа генов циклинов Арабидопсиса. Для множественного выравнивания сиквенирования был использован полный белковый сиквенс 50 генов циклинов. Множественный sequence aligment был использован для конструирования дерева некорневых ближайших соседей, основанного на сиквенсе сходства. Соответствующие гены ID циклинов классифицированы тут и представлены в Таблице 1.1. СYCQ1;1 – новое название, которое мы предложили для CYCJ18. В рисе было найдено минимум 50 циклинов, и хотя 10 классов таких же циклинов (A-, B-, C-, D-, H-, L-, SDS, Q-, T- И Р-типы) найдено в Arabidopsis, циклин F-типа, который обнаружен в рисе, не был идентифицирован ни в Arabidopsis ни в других видах растений. Их взаимоотношения з другими группами циклинов, так же хорошо, как и их функции остаются определенными. Стоит указать, что семейство циклинов растений оказывается намного больше, чем в остальных эукариот. Например, геном Caenorhabditis elegans имеет 34 циклина, тогда как у человека найдено около 22.
|