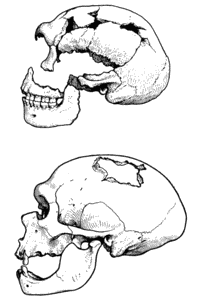
ПРЕДПОСЫЛКИ ОЧЕЛОВЕЧИВАНИЯ1) Большой относительный объем головного мозга, особенно новой коры. У некоторых млекопитающих из китообразных, вследствие большого размера тела, головной мозг также велик, но кора головного мозга развита у них значительно слабее, чем у обезьян. Относительный объем головного мозга человека - 1:43, шимпанзе - 1:61, орангутана - 1:183, гориллы - 1:220, гиббона - 1:73. Нормальная масса мозга современного человека составляет около 1300 г, шимпанзе - 345 г, гориллы - 420 г, орангутанга - 400 г, гиббона - 130 г. У понгид масса мозга может значительно варьировать, достигая у гориллы 750 г. Кора головного мозга человека и человекообразных обезьян имеет большое количество промежуточных нейронов. Чем выше уровень развития центральной нервной системы животного, тем большую роль играют промежуточные нейроны в его высшей нервной деятельности, поскольку они обеспечивают многообразие нервных связей. Считается, что число промежуточных нейронов в каком-либо нервном центре связано прямо пропорциональной зависимостью со способностью данного нервного центра к переработке информации. Некоторые области коры головного мозга человека почти целиком состоят из промежуточных нейронов. Речевая функция человека одновременно связана с такими сенсорными функциями, как слух и зрение, а так же с двигательными функциями рук. Центры, ответственные за эти функции, находятся, в основном, в лобной, затылочной и височной долях мозга. У человека и у человекообразных обезьян эти отделы мозга развиты значительно лучше, чем у остальных млекопитающих. В коре головного мозга приматов (и человека, в частности) есть сенсорные, моторные и ассоциативные зоны. Сенсорные зоны получают информацию от зрительных, слуховых, кинестетических и, в меньшей мере, от других рецепторов. Моторные зоны посылают команды, управляющие движениями. Большую же часть коры головного мозга человека занимают ассоциативные зоны, не имеющие какой-либо явной специализации, но функционально ответственные за объединение и переработку информации. Ассоциативные зоны коры головного мозга очень важны для обеспечения таких высших психических процессов, как память, научение, мышление и речь. 2) Хорошее развитие зрения, слуха и кинестетического чувства и редукция (ослабление) обоняния. Большинство млекопитающих получают информацию об окружающем мире посредством обоняния. Также у большинства млекопитающих зона обонятельной чувствительности мозга доминирует среди остальных сенсорных (чувствительных) зон. Иная картина наблюдается у приматов. У человекообразных обезьян и людей обонятельная зона головного мозга редуцирована до двух маленьких участков – обонятельных луковиц. По сравнению с прочими млекопитающими, пучки осязательных волос на лице понгид редуцированы; у людей этих чувствительных образований нет вовсе. Всё это значит, что сенсорное взаимодействие приматов с окружающей средой происходит, в основном, на основе зрения и слуха. Зрение и слух стали особенно важны тогда, когда предки современных обезьян перешли от обитания в лесах к обитанию на открытых участках африканских саванн. В коре больших полушарий приматов, как в сенсорной, так и в моторной зоне в виде проекции представлены все части тела. Эту функциональную особенность метафорически представляют в виде так называемого «мозгового гомункулюса» – схемы, которую изображают в виде маленького человечка с головой, направленной вниз, и пальцами ног, направленными вверх. Пропорции головы и туловища этого «мозгового гомункулюса» отличаются от таковых пропорций настоящего тела человека, отражая тем самым отличия в неврологическом соответствии различных соматических органов. Площадь поверхности «гомункулюса» пропорциональна сложности управления соответствующими группами мышц настоящего тела человека. Данная метафора показывает, что величина проекций сенсорных и моторных волокон в коре головного мозга несоразмерна с величиной тех участков тела, от которых эти волокна отходят. В головном мозге человека (и человекообразных обезьян) максимальная проекция представлена кистью правой руки (для правшей) и лицевой частью головы. Таким образом, мимика, а также подвижность и чувствительность кистей рук значительно отличают человекообразных обезьян и человека от остальных млекопитающих. Хорошо развитая мимика необходима приматам для обеспечения коммуникативного взаимодействия, передние конечности обеспечивают функции манипулирования и, в конечном итоге, адаптивную функцию преобразования среды обитания. 3) Особое строение передних конечностей. Для обеспечения «предпосылок» трудовой деятельности приматов важны их следующие морфологические особенности: а) пятипалая хватательная конечность с противопоставленным первым пальцем; б) эволюционная замена когтей ногтями; в) способность верхних конечностей к вращательным движениям в локтевом суставе; г) значительный объем движений плеча. Эти морфологические признаки обезьян способствовали становлению рук человека как «трудового органа». Однако, характеризуя функционально-морфологические предпосылки трудовой деятельности, «унаследованные» человеком от обезьян, следует помнить, что главной предпосылкой трудовой деятельности человека является не строение его рук или коры головного мозга, а социально-психологический статус личности, выросшей в человеческом обществе. Под социально-психологическим статусом в данном случае понимаются такие характеристики человека, как идеалы, сформировавшиеся в ряду исторических поколений, воля, ментальность (интеллект, мыслительные механизмы), «творческая жилка», социальная самоидентификация, -- свойства, благодаря которым люди испытывают потребность трудиться, а также перенимают у предыдущих поколений и развивают трудовые навыки и социально нормированные способы трудовых взаимодействий. 4) Длинный период детства приматов и рождение, как правило, одного детеныша за одну беременность. У шимпанзе первая беременность наступает обычно на десятом году жизни, кормление детеныша занимает около 2-3 лет, около матери детеныш держится до 6-8 лет. Удлинение детства увеличивает период формирования адекватного поведения, удлиняет период научения, игр, овладения «нормами жизни» в организованных сообществах биосоциального типа, время освоения «предкультурного» поведения. Под «предкультурным» поведением человекообразных обезьян этологи понимают передачу от поколения к поколению новых приемов поведения, навыков, привычек, традиций, и даже примитивных орудий и умений обращаться с ними. С другой стороны, рождение у приматов одного детеныша за одну беременность является предпосылкой лучшего ухода за ним и его обучения со стороны матери. Детство – это такой период жизни млекопитающего, во время которого он находится под опекой родителей, поэтому добывание пищи и защита от опасности не являются доминантами его поведения в той мере, в которой они присутствуют у взрослых особей. Длительный период детства способствует передаче потомству не только инстинктивного, но также приобретенного (так называемого «выученного») поведения. Зоопсихологи различают облигатное и факультативное научение. Первое представляет собой обязательный минимум, а второе – дополнительное. Было выяснено, что, чем длиннее по времени длится детство у какого-либо вида животных, тем большая доля факультативного научения наблюдается в его поведении. Кроме того, детство – это период игр. Также зоопсихологам известна следующая закономерность: чем «умнее» животное, то есть чем выше у него доля рассудочного поведения, тем больше это животное играет, причем не только в детстве. Играют почти все виды млекопитающих, а среди птиц играют представители группы врановых, являющиеся в этом отряде самыми «сообразительными». В своих играх животные как бы «отшлифовывают» взрослое поведение. Игры способствуют развитию образного мышления даже у животных. У человека, по сравнению с животными, значение игр неизмеримо возрастает. Кроме «естественных» игр, у ребенка появляются социальные игры, имитирующие общественную деятельность взрослых. Кроме практических следствий тренировки какой-либо деятельности, наблюдающейся у животных, игровая способность человека развивает его умственные и социальные навыки. Сюда относится умение распознавать общий контекст событий, происходящих на арене социальных взаимоотношений, и, самое главное, обеспечивает поведенческую динамику подобных событий, быструю смену ведущих контекстов, необходимую для формирования социальной гибкости личности. 5) Утрата сезонности размножения. Способность к социально организованному труду, по мнению многих авторов, является следствием высокой энергетики функционирования тела человека при одновременно высокой экономии энергии на цели размножения. Предпосылки для экономии имеются и у большинства приматов, а именно – утрата сезонности размножения и «приобретение» способности к полицикличности деторождения. Благодаря полицикличности размножения высшие приматы освобождены от власти «половой доминанты». При этом человек, в течение своей взрослой жизни, потребляет значительно больше энергии на единицу массы тела, чем другие млекопитающие. Эту энергию, сэкономив на размножении, человек способен направить на трудовую деятельность. 6) Стадный или групповой образ жизни. Все ныне живущие приматы обладают сложной системой социальной организации. Они живут группами, в которых строго определены как иерархический ранг особи, так и система доминирования. Как правило, доминантом стада обезьян является самец, обладающий сильным и подвижным типом нервной системы. Особь - доминант является образцом для подражания прочих особей. При опасности особи более низкого ранга ориентируют свое поведение на поведение доминанта. Если доминант ведет себя спокойно, значит, опасности нет. Генетические характеристики доминанта передаются по наследсту, поскольку доминант обезьяньего стада имеет преимущества при спаривании, не позволяя самцам более низких рангов приближаться к самкам. Общим свойством любых зоологических иерархий является периодическая смена доминанта, что вызывает «перемешивание» генофонда группы. Интересно, что чаще всего агрессия доминанта направлена на особь - субдоминанта (вторую по рангу). Эти животные находятся в постоянном стрессовом состоянии, а при «смене власти» доминантом в стаде становится не субдоминант, а какая-либо другая особь. Групповой образ жизни животных обеспечивает лучшую защиту от врагов, передачу прогрессивных приобретенных форм поведения, способствует развитию подражательного поведения и коммуникативных навыков, обеспечивает выживание детенышей при гибели биологических родителей, способствует функциональному разделению стада и т.д. 7) Высокий уровень развития высшей нервной деятельности, исследовательской и манипулятивной активности. Шимпанзе широко используют природные предметы, такие, как палки, ветки, камни для добывания пищи и при угрозе и нападении. Свои орудия они могут хранить в специальных местах. Лабораторные исследования над шимпанзе показали, что для них характерна даже элементарная способность к анализу устойчивых причинных связей между явлениями. Некоторые, наиболее сообразительные, обезьяны узнают свой образ при отражении его в зеркале. Шимпанзе обладают образной памятью, элементами абстрактного мышления (они способны к выбору предмета по образцу, к выбору «третьего лишнего» предмета из образцов, предоставленных зоопсихологами), в ходе лабораторного обучения они даже приобретают познавательные навыки. В природных условиях для поведения шимпанзе отмечены кооперативные действия (нападение, защита, помощь и взаимопомощь), они способны организовать коллективную охоту. Также хорошо развиты у шимпанзе способности к подражанию, наблюдению, скрытому научению. Шимпанзе умеют быстро перенимать друг у друга новые приемы поведения, изобретенные ими впервые или увиденные у человека. 11) Сложная внутривидовая коммуникация. Обезьяны, обитая, в основном, в стадах или семейных группах, что встречается значительно реже, обладают сложной мимикой, основное назначение которой – общение с сородичами. С помощью мимики, жестов и специальных положений тела они передают сородичам сигналы о своих намерениях и эмоциональном состоянии. Звуки, издаваемые обезьянами, также служат целям коммуникации внутри стада, широко используются при общении матери и детеныша, половых партнеров. Для них характерно разделение ролей при совместной охоте, в ходе миграции при передвижении стада, при воспитании детенышей и т. д. 12) Функциональная асимметрия тела. Для обезьян, как и для большинства из людей, характерна ориентация и передвижение преимущественно в правую сторону, а также преимущественная праворукость, наблюдаемая в процессе манипуляции. Этот признак обезьян, по-видимому, присутствовал в поведении ископаемых антропоидов и гоминид. Он мог сыграть чрезвычайно значительную роль в ходе формирования функциональной асимметрии коры головного мозга современного человека, складывающейся в его индивидуальном развитии – онтогенезе. ОСНОВНЫЕ СТАДИИ АНТРОПОГЕНЕЗА Считается, что первая популяция человеческих существ имеет древность 2—2,5 млн лет. Но, по мнению многих ученых, первые люди могли появиться значительно раньше. I. Предшественники человека — австралопитековые. Формирование человека как биологического вида проходило через четыре основных этапа эволюции в пределах семьи гоминид: предшественник человека (протоантроп); самый давний человек (архантроп); древний человек (палеоантроп); человек современного типа (неоантроп). НОMO Первым представителем рода Homo считается, с рядом оговорок, вид Homo habilis, или «человек умелый». Он появился в Восточной Африке около 2,4 млн. лет назад. Примерно к этому же времени относятся и древнейшие из известных сейчас каменных орудий со следами намеренного изготовления, знаменующие собой для археологов начало палеолита (древнекаменного века) и начало нашей культурной истории в целом. В ряде случаев эти орудия были найдены вместе с костями человека умелого, чему последний и обязан таким названием. От своих предшественников габилис отличался, прежде всего, формой черепной коробки, приобретшей более плавные и округлые очертания, чем у австралопитеков. Это было вызвано изменениями, которые претерпел мозг. Даже если судить только по величине мозга, не касаясь его строения, различия между австралопитеками и человеком умелым все равно будут впечатляющими. У первых объем эндокрана колеблется от 400 до 500 см3, почти никогда не выходя за эти рамки, а у второго он редко бывает меньше 600 см3 и в отдельных случаях достигает даже 700 с лишним кубических сантиметров. Кроме того, как показывает изучение рельефа внутренней поверхности мозговой полости габилисов, у них уже формировался подобный человеческому узор борозд, и даже появились довольно хорошо выраженные выпуклости в тех областях коры, где у людей находятся речевые центры (поле Брокá и поле Вернике).
Рис. 1.6. Череп гомо габилис из Восточной Африки. Некоторые исследователи считают, что габилисов было бы правильнее разделить на два, а то и на три вида, поскольку между черепами, традиционно относимыми к этой группе гоминид, существуют заметные различия. Например, обладателя знаменитого черепа KNM-ER 1470(рис. 1.6), выделяющегося своим необычайно большим размером, часто зачисляют в особый вид человека рудольфского (Homorudolfensis), а иногда даже определяют как позднего представителя рода кениантропов, с которым мы уже встречались выше. Правы сторонники подобных перетасовок или нет, покажут будущие исследования, но, в любом случае, именно человек умелый или какая-то из его разновидностей кажутся сейчас наиболее вероятными предками всех более поздних членов рода Homo. Другие подходящие на эту роль кандидаты пока просто отсутствуют – то ли еще не найдены, то ли их никогда и не существовало. Наиболее поздние костные останки человека умелого имеют возраст около 1,7 млн. лет. Примерно тогда же появляются и первые представители вида Homoerectus – человек прямоходящий. Эта смена форм гоминид совпадает с важными событиями в истории климата: она приходится на самое начало плейстоцена или, иначе говоря, ледникового периода. К этому времени значительную часть Северного полушария уже покрывали льды, а на примыкавших к ним пространствах господствовали тундры и лесотундры. Изменения коснулись и низкоширотных регионов, но там они были не столь резкими. В Африке, все еще остававшейся единственным местом обитания гоминид, сократилось количество осадков и возросла амплитуда температурных колебаний, но в общем на этом континенте по-прежнему сохранялись сравнительно мягкие климатические условия. Плейстоценовый отрезок эволюции человека, т.е. ее последние полтора с небольшим миллиона лет, можно с большей или меньшей степенью условности разделить на три частично перекрывающихся периода или стадии: стадию архантропов, стадию палеоантропов и стадию неоантропов. Архантропы1— это общее название для гоминид, в прошлом именовавшихся питекантропами и синантропами, а сейчас относимых большинством антропологов к уже упоминавшемуся выше виду гомо эректус (Homo erectus), который пришел на смену габилисам. Иногда этот вид делят на два и его более раннюю африканскую форму называют эргастер (Homo ergaster). Термин палеоантропы может служить в качестве собирательного обозначения всех близких и дальних потомков архантропов, за исключением людей современного физического типа. Как правило, этих потомков зачисляют в виды человек гейдельбергский (Homo heidelbergensis) и неандерталец (Homo neanderthalensis), хотя есть и иные варианты их классификации. Наконец, неоантропы – это, согласно традиционному словоупотреблению, люди современного физического типа, т.е. члены вида гомо сапиенс. Архантропы, судя по находкам на восточном (местонахождение Кооби-Фора) и западном (местонахождение Нариокотоме) побережьях озера Туркана, появились где-то 1,7-1,8 млн. лет назад. Первые полмиллиона лет своего существования они провели, по-видимому, почти безвыходно в Африке, совершая иногда более или менее продолжительные «вылазки» лишь в некоторые соседствующие с ней районы. Одним из таких районов, несомненно, был Ближний Восток, а другим, более отдаленным — Кавказ, где недавно обнаружены хорошо сохранившиеся черепа и нижние челюсти ранних эректусов. По настоящему широкое расселение гоминид за пределы их африканской прародины началось примерно миллион лет назад: начиная с этого времени следы их присутствия (костные останки и каменные орудия) все чаще встречаются в восточной части Азии и на западе Европы. Если отвлечься от ряда мелких деталей, то не будет преувеличением сказать, что для вида гомо эректус в общем характерен уже вполне человеческий скелет, который по многим параметрам, в том числе по размерам и пропорциям, очень близок к современному состоянию. Особенно показательно в этом смысле соотношение длины бедренной, плечевой и предплечевых костей. У шимпанзе оно равняется примерно 1:1, а у людей, хотя плечо имеет почти тот же абсолютный размер, бедро удлинено, а предплечье укорочено. Впервые эти человеческие пропорции фиксируются именно у эректусов около 1,5 млн. лет назад. Вид гомо эректус отличается также от более ранних гоминид возросшими размерами тела и уменьшением различий по этому признаку между полами. Кроме того, уже у самых ранних эректусов (т.е. тех, кого причисляют иногда к виду гомо эргастер) в отличие от габилисов и австралопитеков полностью отсутствуют какие бы то ни было черты, свидетельствующие о сохранении приспособленности к древесному образу жизни.
Рис. 1.7. Череп гомо эректус из Восточной Азии. Средний размер эндокрана у архантропов вида гомо эректус составляет около 1000 см3. Однако, несмотря на увеличение мозга, на черепе еще сохраняется много примитивных черт (рис. 1.7). Лобная кость у эректусов низкая и покатая, с прямо выступающим надглазничным валиком, затылок угловатый, стенки черепа толстые, а наибольшая его ширина приходится не на среднюю часть, как у современных людей и неандертальцев, а на основание, как у австралопитеков. Для лицевого скелета характерно еще заметное выступание вперед ротовой части (хотя и не столь сильное, как у более ранних гоминид), а нижняя челюсть лишена подбородочного выступа и убегает назад. Жевательные зубы крупнее, чем у современных людей, резцы тоже. В то же время, по относительному размеру зубов и нижней челюсти, а также по толщине эмали уже самые ранние архантропы отличаются от современных людей гораздо меньше, чем австралопитеки и габилисы. После широкого расселения гомо эректус за пределы Африки между разбросанными в пространстве популяциями этого вида стало накапливаться все больше и больше различий. Это совершенно естественно. Природные условия Африки, Восточной Азии и Средиземноморья предъявляли к обитавшим в этих регионах гоминидам разные требования, отбор работал в разных направлениях и с разной интенсивностью, и, как следствие этого, эволюционные изменения должны были вести к формированию специфических особенностей у каждого из географических вариантов сначала архантропов, а затем и сменивших их палеоантропов. Конечно, вполне возможно, что процесс расхождения в какой-то мере сглаживался за счет диффузии генов, которые даже при скрещивании только между соседними популяциями со временем могли распространяться на многие тысячи километров, но вряд ли этого было достаточно для поддержания на протяжении сотен тысяч лет биологического единства потомков гомо эректус. Даже если допустить, что они оставались членами одного вида в репродуктивном смысле, т.е. сохраняли способность скрещиваться и давать при этом не стерильное потомство, углубляющиеся различия в анатомии и в поведении, а также географическая изоляция должны были вести к тому, что на практике эта способность реализовывалась нечасто. Подобная ситуация характерна для многих современных животных, например, для волка и собаки. Еще один пример - африканские львы и бенгальские тигры. Внешне столь непохожие и относимые к разным видам рода Panthera, они, тем не менее, не разделены еще бесповоротно барьером репродуктивной изоляции, что успешно доказывают в зоопарках. Эволюционная судьба различных географических популяций архантропов сложилась, вероятно, по-разному. Согласно преобладающей сейчас точке зрения, лишь африканские представители вида гомо эректус могли иметь прямое отношение к происхождению людей современного физического типа. Такие взгляды называют моноцентризмом, поскольку предполагается, что первоначальное становление гомо сапиенс было связано с одним, относительно ограниченным регионом. Противоположную позицию занимают исследователи, отстаивающие так называемую «мультирегиональную», или полицентристскую теорию, согласно которой эволюционное превращение эректусов в гомо сапиенс происходило параллельно и повсеместно, то есть и в Африке, и в Азии, и в Европе, при постоянном обмене генетическим материалом между популяциями, населявшими эти континенты. Хотя спор между моноцентристами и полицентристами, имеющий длительную историю, все еще далек от окончательного завершения, инициатива сейчас явно в руках первых, а вторые занимают оборонительную позицию.
Рис. 1.8. Череп раннего палеоантропа из Южной Африки. Около полумиллиона лет назад или немного раньше в большинстве регионов, освоенных архантропами, им на смену приходят гоминиды несколько иного типа. Являясь потомками гомо эректус и сохраняя еще многие присущие ему черты, они, тем не менее, повсеместно отличаются от своих предшественников общей формой черепа (рис. 1.8) и заметно возросшим объемом мозговой полости. У этих гоминид, как правило, более широкий, чем у эректусов, лоб и более округлый затылок, лишенный свойственного архантропам угловатого выступа. Для них также характерно разрастание теменной области черепа. Размер эндокрана на этой стадии эволюции, которую выше мы уже договорились именовать стадией палеоантропов, обычно превышает 1100 см3, а в отдельных случаях достигает даже размеров, типичных для современных людей (1300 см3 и более). Всех ранних палеоантропов независимо от того, найдены ли они в Южной Африке, в Восточной Азии, или в Западной Европе, сейчас относят обычно к виду человек гейдельбергский, но существуют и более дробные их классификации, придающие большее значение местным особенностям ископаемых материалов из разных регионов. В частности, костные останки древнейших известных нам европейских гоминид настолько своеобразны, что зачастую их выделяют в особый вид, получивший название гомо антецессор (Homo antecessor). Этот вид иногда рассматривают в качестве последнего общего предка неандертальцев и современных людей. Согласно другой точке зрения, он стоит у истоков линии, ведущей к неандертальцам, и не имеет прямого отношения к происхождению гомо сапиенс. Картина, конечно, в итоге получается крайне запутанная, и пройдет, вероятно, еще немало времени, прежде чем она будет прояснена. Среди европейских палеоантропов, следующих по времени за антецессором, довольно отчетливо выделяются две группы. Это, во-первых, так называемые пренеандертальцы, которых почти все исследователи тоже относят теперь к виду человека гейдельбергского, и, во-вторых, произошедшие от них «классические» неандертальцы, выделяемые, как правило, в самостоятельный вид. Пренеандертальцы жили в период примерно от 500 до 150 тыс. лет назад и заселяли юг и запад Европы от Балкан до Британских островов и от Рейна и Дуная до Пиренейского полуострова. Классические неандертальцы появились приблизительно на рубеже среднего и позднего плейстоцена, т.е. 130—150 тыс. лет назад. Некоторые антропологи, впрочем, допускают, что в качестве наиболее древних представителей неандертальского вида следует, возможно, рассматривать уже гоминид из Сванскомба в Англии и Сима де лос Хэсос в Испании, живших 300—400 тыс. лет назад. Местом происхождения неандертальцев, наверняка, была Европа, но впоследствии они значительно расширили ареал своего обитания, заселив сначала Западную Азию, а затем и более отдаленные районы на востоке вплоть до южной Сибири. Этот вид существовал на протяжении, как минимум, ста тысячелетий: последние его представители жили в ряде районов Евразии еще 30 тыс. лет назад и даже, возможно, несколько позднее. Считается, что особенности строения неандертальского скелета во многом обусловлены необходимостью приспособления к холодному климату Европы в периоды позднечетвертичных оледенений, когда средняя температура была значительно ниже современной, а южная граница зоны тундр проходила там, где сейчас простираются лесостепи. Адаптацией к холоду объясняют, например, такие признаки, свойственные неандертальцам, как общая массивность тела, сравнительно большая толщина трубчатых костей, укороченные пропорции конечностей, а также специфическое строение носового отверстия – более широкого, чем у современных людей. Массивность тела, имеющая следствием уменьшение площади его поверхности, приходящейся на единицу объема, помогала сэкономить тепло и энергию (поэтому у широко расселенных видов млекопитающих представители северных популяций, как правило, крупнее своих южных сородичей – так называемое правило Бергмана). Крупный нос также играл адаптивную роль, предотвращая попадание слишком холодного воздуха в легкие при дыхании. По всем перечисленным и некоторым другим особенностям неандертальцы отличались в равной мере и от гомо сапиенс, и от более древних гоминид, чья эволюционная история протекала в основном в низких широтах с их теплым климатом.
Рис. 1.9. Черепа неандертальцев из Западной Европы. При относительно невысоком среднем росте (165—170 см) для неандертальцев характерна массивность костей скелета, свидетельствующая о большой мышечной массе. Они, несомненно, обладали значительной физической силой, но красавцами их, если руководствоваться нынешними эстетическими мерками, назвать было бы трудно. Впрочем, иной неандерталец, одетый в современный костюм, пожалуй, вполне органично смотрелся бы в современной толпе и его появление в ней не вызвало бы особого ажиотажа. Свой низкий и покатый лоб он мог бы спрятать под шляпой, а крупные широко расставленные орбиты глаз с развитыми надбровными дугами скрыть за темными очками. Коренные зубы у неандертальцев были меньше, чем у других гоминид, за исключением гомо сапиенс, а резцы, наоборот, крупнее, чем у предшественников и гораздо крупнее, чем у людей современного физического типа. Черепная коробка у них длинная и широкая (рис. 1.9). Специфическими чертами анатомии черепа, отличающими неандертальцев от всех современных им и более поздних гоминид, считаются его субсфероидная форма (если смотреть сзади), заметное выступание вперед нижней части лица, наличие затылочного валика и жевательного гребня за слуховым отверстием. Еще одним признаком, имеющимся у почти всех неандертальцев и отсутствующим у большинства представителей других видов рода Homo, является так называемый «ретромолярный пробел» — пустое пространство между задними коренными зубами и восходящей ветвью нижней челюсти. Несмотря на общий «архаизм» облика неандертальцев, абсолютная величина мозговой полости у них и в среднем, и по размаху крайних значений (1200—1600 см3) практически не отличалась от таковой у современных людей, даже несколько превосходя ее. На этом основании в 60-е-80-е годы прошлого века обе группы часто рассматривали как два подвида одного вида, но впоследствии такой подход к их классификации утратил популярность. Сопоставление ДНК, которую удалось извлечь из нескольких неандертальских костей, с ДНК современных людей показало, что генетическое расстояние между этими формами гоминид в три с лишним раза превышает расстояние между ныне существующими расовыми группами человека. По строению скелета и черепа неандерталец и гомо сапиенс тоже различаются между собой больше, чем следовало было бы ожидать от представителей одного биологического вида. Руководствуясь этими фактами, большинство антропологов сейчас рассматривает их как два разных вида. К началу верхнего плейстоцена, т.е. как раз к тому времени, когда в Европе появляются неандертальцы, в Африке заканчивается в основном сложение комплекса анатомических признаков, характеризующих людей современного физического типа. Об этом свидетельствуют многочисленные находки костей древнейших гомо сапиенс, сделанные на юге и востоке континента. Некоторые из них имеют возраст свыше ста тысяч лет, а совсем недавно на местонахождении Херто в Эфиопии обнаружили сразу три черепа, очень похожих на современные, но относящихся ко времени около 160 тысяч лет назад. Один из этих черепов сохранился почти целиком. Предшественниками гомо сапиенс на африканском континенте и, скорее всего, их прямыми предками были местные палеоантропы, которых обычно относят вместе с их европейскими и азиатскими современниками к виду человека гейдельбергского, но иногда выделяют в особые виды гомо родезиенс, или человек родезийский (Homorhodesiensis), и гомо хельмей (Homohelmei). В первый из них при этом включают ранних африканских палеоантропов, а во второй поздних. Если уклонение линии, ведущей к неандертальцам, от предкового ствола можно, по крайней мере, частично объяснить приспособлением к природным условиям севера, то формирование большинства признаков, выделяющих людей современного физического типа, остается загадкой. Неизвестно, в каком именно районе Африки появился впервые новый вид человечества, в каких условиях происходило его становление, какие требования среды обусловили давление отбора в соответствующем направлении. Неизвестно, впрочем, и был ли это действительно новый вид в полном смысле слова, или только особый анатомический тип гоминид, отличающихся от своих предшественников и современников определенным комплексом анатомических и поведенческих особенностей, но не отделенных от них барьером репродуктивной изоляции. Даже дать четкое определение того, какие именно черты в строении черепа и скелета отделяют людей современного физического типа от всех остальных представителей рода Homo, по мнению многих антропологов, гораздо труднее, чем очертить, скажем, круг специфических неандертальских признаков. В числе наиболее заметных особенностей, свойственных гомо сапиенс, округлая форма затылка, вертикальный лоб, относительно плоское лицо, слабая, как правило, выраженность надбровных дуг, наличие подбородочного выступа (рис. 1.10).
Рис. 1.10. Череп верхнепалеолитического гомо сапиенс из Западной Европы. Судя по археологическим и палеоантропологическим находкам, широкое распространение гомо сапиенс за пределы Африки и Ближнего Востока началось не раньше 50—60 тыс. лет назад. Сначала, по всей видимости, ими были колонизированы южные, тропические и субтропические районы Азии и Австралия, близкие по своему климату и ландшафтам к тому региону, где совершалось первоначальное становление нашего биологического вида. Несколько позже, в период от 35 до 40 тыс. лет назад, люди современного физического типа впервые появились в Европе. Здесь, напротив, им пришлось приспосабливаться уже к совершенно непривычным природным условиям, намного более суровым и переменчивым, чем в тех областях к югу от Сахары, откуда вышли когда-то их предки. В Европе пришельцы столкнулись с довольно многочисленным коренным населением, т.е. с неандертальцами, которых, вероятно, уже встречали раньше на Ближнем Востоке и в Передней Азии. Результатом очередной встречи для неандертальцев стало полное вымирание, но оно было не одномоментным, а растянулось на многие тысячелетия. Лишь 30 тыс. лет назад или даже несколько позже людьми современного физического типа были колонизированы юго-запад Пиренейского полуострова, а также Крым и некоторые другие районы, остававшиеся до той поры последним прибежищем ев ропейских неандертальцев. Происходило ли в ходе колонизации смешение коренного и пришлого населения - неясно. Упоминавшиеся уже результаты сопоставления ДНК из костей неандертальцев с ДНК современных людей предполагают скорее отрицательный, чем положительный, ответ на такой вопрос, но работы в этой области только начинаются, и было бы преждевременно, основываясь на них, делать уже сейчас сколько-нибудь далеко идущие выводы. К тому же, вопреки результатам палеогенетических исследований, ряд черт в анатомии отдельных частей скелета поздних неандертальцев и ранних европейских гомо сапиенс указывает на возможность того, что их сосуществование все же не обошлось без некоторого, пусть и незначительного, обмена генами. К концу ледникового периода, или, точнее говоря, к началу современного межледниковья, именуемого голоценом, ойкумена гомо сапиенс приобрела, практически, свои современные очертания. Это произошло после того, как примерно 15 тыс. лет назад началось заселение Америки. Правда, некоторые исследователи считают, что последнее событие произошло гораздо раньше, но убедительные доказательства в пользу этого по
|