
Механизм мышечного сокращения.
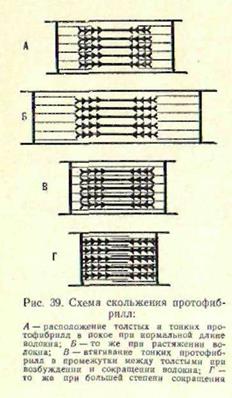
Когда мышца находится в несокращенном (расслабленном) состоянии, нити актина и миозина лишь частично продвинуты относительно друг друга, причем каждой нити миозина противостоят, окружая ее, несколько нитей актина. Сокращение мышечного волокна, а, следовательно, и мышцы в целом связано с тем, что тонкие актиновые протофибриллы втягиваются в глубь промежутков между миозиновыми. Эта теория по поводу механизма мышечного сокращения, основанная на современных электронно - микроскопических данных, получила название теории скольжения: актиновые нити, втягиваясь в глубь анизотропных дисков, как бы скользят между миозиновыми. Под электронным микроскопом можно рассмотреть, что при сокращении мышцы ширина анизотропных (темных) дисков не меняется, а ширина изотропных (светлых) уменьшается. При значительном сокращении эти диски совсем исчезают — полностью втягиваются внутрь анизотропных (рис.2). Иннервация скелетной мышцы. Мышца сокращается лишь при возникновении в ней возбуждения — после того как произойдет целый ряд процессов (электрических, химических), обеспечивающих возможность выполнения ею специфической функции — укорочения, или развития напряжения. А возбуждается мышца рефлекторным путем —под влиянием импульсов, которые передаются к ней от центральной нервной системы по эфферентным (т. е. ценробежным нервам.) В организации двигательных актов участвуют различные отделы центральной нервной системы, но непосредственную связь со скелетными мышцами имеют низшие ее отделы. В так называемых передних рогах спинного мозга расположены тела мотонейронов, длинные отростки которых — аксоны направляются к мышцам туловища и конечностей и заканчиваются в них своими разветвлениями (рис. 3). Аксоны нервных клеток образуют те нервные волокна, посредством которых периферические органы связаны с центральной нервной системой. Через мотонейроны и их аксоны на мышцы оказывают влияние и вышележащие отделы центральной нервной системы — различные отделы головного мозга.
 Помимо эфферентных (двигательных) нервов мышцу иннервируют и афферентные, или чувствительные, нервы. Их окончания связаны с чувствительными образованиям — проприорецепторами. Последние возбуждаются при изменениях состояния мышечных волокон — их сокращении и растяжении. Импульсы от них передаются афферентными нервами в центральную нервную систему и информируют, таким образом, соответствующие нервные центры о длине мышцы и развиваемом ею напряжении. Помимо эфферентных (двигательных) нервов мышцу иннервируют и афферентные, или чувствительные, нервы. Их окончания связаны с чувствительными образованиям — проприорецепторами. Последние возбуждаются при изменениях состояния мышечных волокон — их сокращении и растяжении. Импульсы от них передаются афферентными нервами в центральную нервную систему и информируют, таким образом, соответствующие нервные центры о длине мышцы и развиваемом ею напряжении.
Двигательная единица. иннервирует, и вся эта группа волокон работает как единое целое. В связи с этим мотонейрон и иннервируемая им группа мышечных волокон получили название двигательной единицы (рис.5). Мышечное волокно можно назвать структурной единицей мышцы, а двигательная единица является ее функциональной единицей.
Возбуждение нерва и мышцы. Из рассмотренных выше положений ясно, что мышца сокращается лишь тогда, когда в ней возникнет возбуждение, что импульсы, вызывающие в ней этот процесс, идут из центральной нервной системы, что непосредственной «станцией отправления» являются мотонейроны, по длинным отросткам которых — эфферентным двигательным нервным волокнам — эти импульсы проводятся к мышечным волокнам и передаются через нервно-мышечные синапсы. Следует выяснить более детально, в чем заключается процесс возбуждения. Возбуждение — это очень сложный биологический процесс, который лежит в основе деятельности органов, тканей и клеток организм.а. Возбуждаясь, каждый орган выполняет свою специфическую функцию: например, железы пищеварительного тракта вырабатывают ферменты, железы внутренней секреции-—гормоны, мышцы производят сократительный акт. Наряду с такими совершенно различными специфическими реакциями имеются и общие черты в возбуждении различных органов. Это относится прежде всего к электрическим явлениям — первым и основным процессам, в которых проявляется возбуждение. Электрические свойства нервных и мышечных клеток в состоянии покоя. Мембранный потенциал. В покое клетка имеет определенный электрический заряд. Снаружи сарколемма заряжена положительно, а изнутри — отрицательно. Возникновение этого двойного электрического заряда связано с особыми свойствами мембраны. Она обладает избирательной проницаемостью для различных ионов. Так, она относительно легко пропускает положительно заряженные ионы (катионы) калия (К+) и почти не пропускает катионы натрия (Nа+). Не могут пройти через мембрану и крупные молекулы белковых анионов. Если бы она была проницаема для всех этих веществ, то содержание их внутри и вне клетки стало бы одинаковым. В связи с избирательной и ограниченной проницаемостью мембраны поддерживается разная концентрация различных ионов внутри клетки и в окружающей ее среде — в межтканевой жидкости. Калия внутри клетки содержится в 30—40 раз больше, чем снаружи, а натрия —в 10—12 раз меньше. В силу разности концентраций катионы К+ выходят из клетки наружу (мембрана для них проницаема), ионы же Nа+ проникнуть внутрь не могут (мембрана в состоянии покоя для них почти непроницаема). Не могут выйти наружу через мембрану и анионы. В связи с диффузией К+ наружу и под влиянием электростатических сил притяжения между противоположно заряженными ионами анионы концентрируются у поверхности мембраны изнутри, а катионы — снаружи, образуя, таким образом, на мембране двойной электрический слой, т. е. поляризуя ее. Разность потенциалов между наружной и внутренней поверхностями мембраны, называемая потенциалом покоя (ПП), или мембранным потенциалом (МП), равна 70—90 мв. Изменения электрического состояния клетки при возбуждении. Потенциал действия. При раздражении клетки происходит изменение мембранного потенциала покоя. Возбужденный участок мембраны оказывается снаружи заряженным отрицательно по отношению к своей внутренней поверхности. Иными словами, происходит перезарядка мембраны, смена знаков зарядов — инверсия потенциала покоя. Этот процесс обусловлен изменениями проницаемости мембраны под влиянием раздражения. Она на какое-то короткое время становится значительно более проницаемой для ионов Na+, чем для ионов К+. Ионы Na+, которых в межтканевой жидкости, как уже указывалось, в 10—12 раз больше, чем внутри клетки, начинают проникать внутрь. Нейтрализуя отрицательный заряд внутренней поверхно-сти мембраны в раздражаемом участке, они снижают тем самым существовавшую в покое разность потенциалов, т. е. приводят к деполяризации мембраны. Интересно, что этот процесс сам себя подкрепляет: начинающаяся деполяризация повышает проницаемость мембраны для ионов Nа+; диффузия катионов Na+ углубляет депо-ляризацию; в связи с этим проницаемость для этих ионов становится еще более значительной и т. д. В результате этого процесса происходит не только деполяризация мембраны, но и ее перезарядка: внутренняя поверхность ее в раздражаемом участке становится заряженной положительно, а наружная по отношению к ней — отрицательно. При измерении разности потенциалов между наружной и внутренней поверхностями клетки оказывается, что вместо заряда — 90 мв внутри, который отмечался в состоянии покоя, там обнаруживается заряд + 30— 40 мв. Проникновение положительно заряженных ионов Na+ внутрь клетки привело к развитию электрического процесса, характеризующегося напряжением 120—130 мв {от—90 до +30 мв). Этот процесс — колебание потенциала покоя — получил название потенциала действия (ПД). Он характеризует возникновение возбуждения в нервной или мышечной клетке.
 (рис. 6). Фаза реполяризации начинается в тот момент, когда потенциал действия достигает своей вершины — пика, т. е. амплитуды 120—130 мв. Проницаемость мембраны для ионов Nа+ при этом резко снижается, и дальнейшее их поступление внутрь почти прекращается. В этот момент проницаемость мембраны для ионов К+ оказывается значительно выше. В связи с отмечавшейся выше разностью концентрации калия внутри и снаружи клетки эти катионы начинают интенсивно покидать клетку. К этому процессу присоединяется и извлечение проникших в клетку ионов Nа+ — как бы «выкачивание» их. Все это приводит к восстановлению исходного состояния клетки — положительному заряду наружной поверхности мембраны и отрицательному — внутренней. Фаза деполяризации потенциала действия (восходящая фаза — см. рис. 6) длится около 1 мсек., в некоторых клетках — 0,5 мсек.; фаза реполяризации (нисходящая фаза) значительно длительнее первой. (рис. 6). Фаза реполяризации начинается в тот момент, когда потенциал действия достигает своей вершины — пика, т. е. амплитуды 120—130 мв. Проницаемость мембраны для ионов Nа+ при этом резко снижается, и дальнейшее их поступление внутрь почти прекращается. В этот момент проницаемость мембраны для ионов К+ оказывается значительно выше. В связи с отмечавшейся выше разностью концентрации калия внутри и снаружи клетки эти катионы начинают интенсивно покидать клетку. К этому процессу присоединяется и извлечение проникших в клетку ионов Nа+ — как бы «выкачивание» их. Все это приводит к восстановлению исходного состояния клетки — положительному заряду наружной поверхности мембраны и отрицательному — внутренней. Фаза деполяризации потенциала действия (восходящая фаза — см. рис. 6) длится около 1 мсек., в некоторых клетках — 0,5 мсек.; фаза реполяризации (нисходящая фаза) значительно длительнее первой.
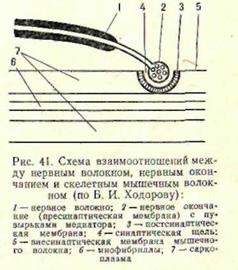
Проведение возбуждения по нервным и мышечным волокнам. С потенциалом действия связано проведение возбуждения по нервным и мышечным волокнам. При возникновении потенциала действия между возбужденным участком и соседними, находящимися в состоянии покоя, возникает разность потенциалов. Наружная поверхность возбужденного участка мембраны, как отмечалось выше, оказывается отрицательно заряженной, а соседнего с ним — находящегося в состоянии покоя — положительно заряженной. В связи с разностью потенциалов между этими соседними участками возникает электрический ток — так называемый местный ток действия. Этот ток является раздражителем участка волокна, находившегося до этого времени в состоянии покоя. Под влиянием раздражения в этом участке начинаются описанные выше процессы — деполяризация, повышение натриевой проницаемости и т. д., т. е. возникает потенциал действия. Затем возбуждается следующий участок волокна и т. д. Таким образом, проведение импульса заключается, по существу, в последовательном, один за другим, возбуждении участков волокна. Передача возбуждения через синапсы происходит, как уже говорилось выше, через посредство химических веществ — медиаторов, вырабатываемых концевыми веточками аксонов. Химическим путем передается возбуждение как в синапсах центральной нервной системы, где разветвления одних аксонов образуют синапсы на теле и дендритах других, так и в нервномышечном, или мионевральном, синапсе. Медиатором окончаний мотонейрона в мышечных волокнах является ацетилхолин. При возбуждении нервных окончаний синаптические пузырьки, в которых содержится медиатор, лопаются, ацетилхолин проникает через пресинаптическую мембрану в синаптическую. щель и вызывает возбуждение постсинаптической мембраны. Последняя обладает высокой чувствительностью к этому воздействию. Под влиянием ацетилхолина повышается ее проницаемость для ионов Na+ и К+, происходит деполяризация и возникает постсинаптический потенциал. С постсинаптической мембраны возбуждение передается на другие (внесинаптические) участки мембраны мышечного волокна снова электрическим путем (см. рис. 4). Значение функционального состояния нервно-мышечного аппарата для развития процесса возбуждения. Возбудимость. Способность живой ткани развивать возбуждение в ответ на раздражение называется возбудимостью. Таким образом, возбудимость является одним из основных свойств живой ткани, обеспечивающих взаимодействие организма со средой. Разные ткани обладают различной возбудимостью. Уровень возбудимости одной и той же ткани тоже изменчив. Умеренные воздействия на ткань повышают ее возбудимость, чрезмерные по силе или длительности понижают. Так, под влиянием разминки возбудимость центральной нервной, системы и нервно-мышечного аппарата повышается, при утомительной работе понижается. Изменение возбудимости происходит закономерно во время протекания каждой волны возбуждения. Когда возникает потенциал действия (в течение фазы деполяризации), ткань становится невозбудимой: она не способна ответить на новое раздражение. Это так называемая абсолютная рефракторная фаза. Постепенно возбудимость ткани восстанавливается до исходного уровня, а затем становится на некоторое время даже выше его. Возбудимость можно измерять. Чем выше возбудимость ткани, тем легче вызвать ее возбуждение — ответную реакцию. Минимальная сила раздражения, которая нужна, чтобы вызвать возбуждение ткани, характеризует так называемый порог возбудимости данной ткани и называется пороговой силой. Уровень возбудимости служит важным показателем функционального состояния ткани. Функциональная подвижность (лабильность). Одним из важных факторов, от которых зависит деятельность возбудимых тканей (таких, как нервные клетки, синапсы, нервно-мышечный аппарат), является скорость протекания возбуждения, получившая название лабильности (Н. Е. Введенский). В одних образованиях волна возбуждения развивается и затухает с большой скоростью, в других значительно медленнее. От скорости возбуждения зависит частота импульсов, которую ткань может развить в единицу времени. Наиболее высока лабильность нервных волокон, значительно ниже лабильность мышечных волокон, нервных клеток и особенно синапсов. Лабильность, как и возбудимость ткани, не постоянна. Умеренные воздействия увеличивают скорость протекания волны возбуждения, чрезмерные — ее уменьшают. Под влиянием разминки, например, лабильность центрально-нервных образований и нервно-мышечного аппарата повышается, при утомлении понижается. Об уровне лабильности можно судить по разным показателям. Н.Е.Введенский предложил измерять ее максимальным числом волн возбуждения, которое может возникнуть в ткани в единицу времени (в 1 сек). Биохимические процессы в мышце при возбуждении. Электрические проявления возбуждения, т. е. потенциалы действия, возникающие на мембране мышечного волокна, приводят к целому ряду химических процессов, которые завершаются механической реакцией волокна — сокращением. Связь между электрическими процессами, происходящими на мембране, и механической реакцией миофибрилл обеспечивается при посредстве ионов кальция (Са++). В состоянии покоя эти ионы находятся преимущественно внутри системы трубочек и полостей, которая пронизывает волокно вдоль (между миофибриллами) и поперек (между отдельными частями —саркомерами миофибрилл). Эти трубочки и полости тоже имеют свои полупроницаемые мембраны, через которые ионы Са++ в покое почти не проникают. Когда возбуждается мембрана мышечного волокна, ее потенциалы действия вызывают деполяризацию мембран трубочек и полостей и повышают их проницаемость. Ионы Са++ выходят (в связи с тем что концентрация их внутри этой системы значительно выше, чем снаружи) и оказываются очень близко от миофибрилл. Ионы Са++ влияют на белок миозин. При рассмотрении строения миофибрилл мышечного волокна уже отмечалось, что они состоят из протофибрилл — тонких (актиновых) и относительно более толстых (миозиновых), чередующихся между собой в поперечном направлении. Миозин, как оказалось, является не только сократительным белком мышцы, а обладает еще и свойствами фермента. Он способен расщеплять очень богатое энергией вещество — аденозинтрифосфорную кислоту (АТФ). Когда волокно находится в состоянии покоя, миозин как фермент неактивен. При воздействии ионов Са++ ферментативные свойства миозина активизируются, и он начинает расщеплять АТФ. За счет химической энергии, которая освобождается при этом, происходит сокращение миофибрилл, т. е. втягивание (скольжение) актиновых протофибрилл в промежутки между миозиновыми. Расслабление мышечного волокна связано с удалением Са++ от сократительного аппарата. Специальные исследования показали, что ионы Са++ после воздействия на миозин, которое привело к расщеплению АТФ и сокращению волокна, как бы «выкачиваются» из сферы сократительного аппарата в ту систему, где они находились до возбуждения мышечного волокна. Работа мышц. В процессе мышечного сокращения потенциальная химическая энергия переходит в потенциальную механическую энергию напряжения и кинетическую энергию движения. Различают внутреннюю и внешнюю работу. Внутренняя работа связана с трением в мышечном волокне при его сокращении. Внешняя работа проявляется при перемещении собственного тела, груза, отдельных частей организма (динамическая работа) в пространстве. Она характеризуется коэффициентом полезного действия (КПД) мышечной системы, т.е. отношением производимой работы к общим энергетическим затратам (для мышц человека кпд составляет 15—20%, у физически развитых тренированных людей этот показатель несколько выше).
|
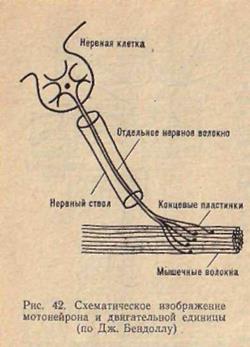 Как уже указывалось выше, аксон мотонейрона, подходя к мышце, ветвится, образуя множество окончаний. Каждое из этих окончаний образует синапс на одном мышечном волокне. Таким образом, каждый мотонейрон снабжает своими окончаниями целую группу мышечных волокон. При возбуждении мотонейрона возбуждаются все мышечные волокна, которые он
Как уже указывалось выше, аксон мотонейрона, подходя к мышце, ветвится, образуя множество окончаний. Каждое из этих окончаний образует синапс на одном мышечном волокне. Таким образом, каждый мотонейрон снабжает своими окончаниями целую группу мышечных волокон. При возбуждении мотонейрона возбуждаются все мышечные волокна, которые он


