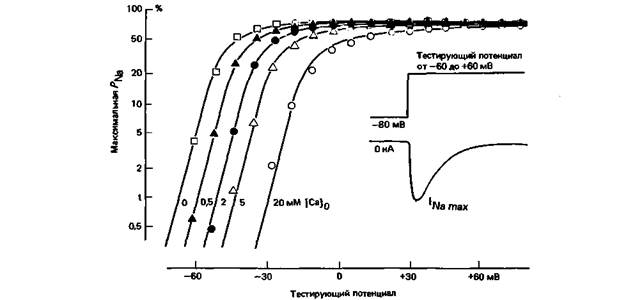
Ионные токи во время следовых потенциаловВо многих клетках после быстрой деполяризации, которая соответствует потенциалу действия, развиваются де- или гиперполяризационные следовые потенциалы (рис. 2.4 и 2.5). Природа таких следовых потенциалов может быть разной; кратко опишем два наиболее важных типа. Кратковременный гиперполяризационный следовой потенциал наблюдается сразу после реполяризации во многих нервных клетках и в некоторых клетках миокарда (рис. 2.5). Этот следовой потенциал представляет собой избыточную реполяризацию; когда во время фазы реполяризации потенциал достигает уровня покоя, gK еще не возвращается к своему исходному уровню (рис. 2.7), т. е. gK в отличие от gNa превышает уровень покоя. Следовательно, мембранный потенциал ближе подходит к значению Ек, чем это было в покое. Возникающая в результате гиперполяризация затухает по мере спада повышенного уровня gK. Такой механизм кратковременной гиперполяризации после потенциала действия участвует в развитии ритмического возбуждения; мы вернемся к нему ниже (с. 48). Длительные гиперполяризационные следовые потенциалы,которые суммируются при достаточно высокой частоте возбуждения, особенно хорошо выражены в тонких нервных волокнах позвоночных-волокнах группы IV. Такие длительные гиперполяризационные следовые потенциалы обеспечиваются электрогенным Na+-насосом (с. 14-17), который удаляет из клетки Na+, вошедший в нее во время возбуждения [28]. Они исчезают, если предотвратить активность насоса веществами, блокирующими метаболизм, например ДНФ (ср. рис. 1.7). «Стабилизирующее» влияние [Са2+]0 на потенциал покоя. Зависимость Na+-токов от тестирующего потенциала, которую иллюстрирует рис. 2.6, может изменяться при различных воздействиях. Блокада некоторых Na+-каналов тетродотоксином или аналогично действующими веществами, а также изменения плотности Na+-каналов в мембране влияют только на амплитуду, но не на зависимость от потенциала или временной ход Na+-токов. Характерные смещения зависимости мембранных токов от потенциала происходят при изменениях внеклеточной концентрации Са2+, [Са2+]0. На рис. 2.10 графики зависимости максимальной Na+-проницаемости, PNa, от тестирующих потенциалов (абсцисса) построены при разных значениях [Ca2 + ]0. Графики для PNa, построенные в логарифмическом масштабе, вначале имеют вид поднимающейся прямой, а затем выходят на уровень насыщения. Влияние [Са2+]0 заключается в параллельном смещении графиков зависимости PNa от потенциала вдоль оси абсцисс: при [Са2+]0 = 0 небольшие деполяризации вызывают значительные изменения PNa, тогда как при высоких [Са2+]0 такой же Na+-ток может возникнуть только при деполяризации не менее чем на 35 мВ. Следовательно, снижение [Са2 + ]0 способствует генерированию потенциалов действия во время деполяризации. Влияние [Ca2+]0 на PNa, показанное на рис. 2.10, осложняется еще одним эффектом, действующим в том же направлении,- влиянием на зависимость инактивации от потенциала. Кривые зависимости INa max от исходного потенциала (рис. 2.8) при изменениях [Ca2 + ]0 смещаются вдоль оси абсцисс точно таким же образом, как и кривые на рис. 2.10. В результате снижение [Са2 + ]0 сопровождается не только более значительным увеличением PNa в ответ на такую же деполяризацию (рис. 2.10), но и абсолютным уменьшением максимально возможного прироста INa (рис. 2.8). Общий эффект, однако, состоит в том, что уменьшение [Са2+]0ведет к снижению порога генерации потенциала действия, т.е. повышает возбудимость, тогда как увеличение [Са2 + ]0 «стабилизирует» мембранный потенциал. Заметные локальные сдвиги [Ca2 + ]0 не так уж необычны в организме; например, в ЦНС при усилении активности (особенно в синапсах; см. с. 62) поступление Са2 + в клетки ведет к снижению [Ca2 + ]0 в ограниченных межклеточных пространствах (рис. 2.3); возбудимость клеток повышается, что может сопровождаться генерированием разрядов судорожного типа [13]. Общее снижение [Са2 + ]0 в плазме крови вызывает синдром тетании,при котором нерегулируемое возбуждение мышц приводит к судорогам. Удивительное параллельное смещение графиков потенциалзависимости Na+-токов (и других мембранных токов) при повышении [Са2 + ]0 интересно с физической точки зрения. Как показывает представленная на рис. 2.15 модель Na+-канала, наружная сторона мембраны несет фиксированные заряды, главным образом отрицательные. Они принадлежат фосфо- и гликолипидам, а также гликопротеинам (рис. 1.2). Эти заряды удерживают около мембраны ионную оболочку, которая, по приблизительной оценке, обеспечивает примерно половину общего градиента мембранного потенциала, так что канальные белки «чувствуют» не более половины разности потенциалов, существующей между внутренней средой и наружной поверхностью клетки [3, 26]. Ионы Ca2+ взаимодействуют с фиксированными зарядами на поверхности плазматической мембраны, нейтрализуя их. Поэтому при высокой [Са2 + ]0 общий наружный отрицательный заряд снижается, а отрицательный потенциал, действующий на ионные каналы, увеличивается. По этой причине для получения при 20 мМ [Са2 + ]0 такого же повышения PNa, что и при 2 мМ [Ca2 ]0, потребуется примерно на 20 мВ более значительная деполяризация. И наоборот, при снижении [Са2+]0 отрицательный поверхностный заряд возрастает и графики потенциалзависимости смещаются в сторону меньших значений деполяризации. Эффекты поверхностных отрицательных зарядов рассматривались здесь не только для того, чтобы объяснить влияние изменений [Са2 + ]0. Смещения потенциалзависимости, аналогичные представленным на рис. 2.10,наблюдаются и при сдвигах внеклеточного pH. При снижении pH возрастает [Н+]о, что уменьшает отрицательный заряд поверхности мембраны-эффект, аналогичный результату повышения [Са2+]0. Снижение pH до 4,5, так же как повышение [Са2 + ]0 на рис. 2.10, может вызвать смещение активации PNa на 25 мВ. Изменения pH тканей в зависимости от метаболизма вполне возможны. Состояние поверхностного заряда мембраны может также влиять на связывание и активность ионизированных веществ, действие которых аналогичным образом зависит от [Са2+]0 и от pH [3, 26, 33]. ГЛАВА 2. ПЕРЕДАЧА ИНФОРМАЦИИ ПОСРЕДСТВОМ ВОЗБУЖДЕНИЯ 35
|