
ИССЛЕДОВАНИЕ МЕМБРАННОГО ПОТЕНЦИАЛА КЛЕТОК СИМПЛАСТА ВЫСШЕГО РАСТЕНИЯВ любом многоклеточном организме клетка представляет собой его основную структурную и функциональную единицу. Плазматическая мембрана одной клетки - пограничное образование между клеткой и внешней средой — в многоклеточном организме трансформируется в многоклеточное образование, которое представляет собой симпласт в организме высшего растения и базальную систему в организме млекопитающего. Симпласт высшего растения - это пласт клеток, связанных между собой низкоомными контактами, управляемыми мембранными потенциалами плазматических мембран клеток, его составляющих. Плазматические мембраны клеток симпласта образуют непрерывное двухслойное образование, разделенное внутриклеточным пространством составляющих его клеток. Несмотря на тесное объединение клеток, они не теряют в симпласте своей индивидуальности, что и определяет существование разности электрических потенциалов между клетками симпласта и базальной системы. В связи с этим логично предположение, что мембранный потенциал - регулятор процессов жизнедеятельности одной клетки выполняет те же функции в симпластном образовании многоклеточного организма. По симпластному образованию (аналог нервного волокна) распространяется потенциал деполяризации в ответ на его повреждение, превышающее некоторый порог. В связи с неоднородностью клеток симпласта по их морфологическим и физиологическим свойствам и существованию из-за этого разности электрических потенциалов по симпласту распространяется не потенциал действия, а вариабельный потенциал. Он назван вариабельным Хоувинком в 1935 г. из-за вариабельности его формы и амплитуды. От потенциала действия он отличается, главным образом, пологой вершиной и большой длительностью заднего фронта, которая часто составляет десятки минут. Повреждение симпласта в любой его области, если оно превышает порог, приводит к распространению вариабельного потенциала по симпласту и его переходу в новое функциональное состояние. Таким образом, симпласт высшего растения по своим структурным особенностям удовлетворяет требованиям, предъявляемым к однопараметрической системе регуляции процессов жизнедеятельности посредством мембранного потенциала, который существует в каждой структурной единице системы и его надпороговое изменение в любой области системы приводит к распространению изменения и переходу всей системы на новый уровень регуляции. Для выяснения принципов функционирования электрической системы регуляции необходимо найти связь между разностями электрических потенциалов по симпласту в покое и характеристиками вариабельного потенциала повреждения. Электрофизиологические характеристики растений исследуются методом внеклеточного отведения потенциалов между областями, расположенными на поверхности стебля относительно корня. Постоянное электрическое поле (ПЭП) измеряется неполяризующимися (каломельными или хлорсеребряными) электродами, соединенными с поверхностью объекта электролитическими контактами. Разность электрических потенциалов (РЭП) между стеблем и корнем существует у всех растений, как и распределение РЭП вдоль по стеблю. Распределение разностей электрических потенциалов по стеблю относительно корня образует постоянное электрическое поле растения. Существование ПЭП обусловлено рядом причин: 1. Клетки, составляющие симпласт, неоднородны по функциональной роли и метаболической активности и поэтому обладают разными мембранными потенциалами. 2. Между клетками существуют высокопроницаемые контакты переменной проницаемости, которая регулируется мембранными потенциалами этих клеток. 3. Симпласт высших растений структурно неоднороден - имеются ветвления, пересечения и изменения диаметров проводящих пучков. Из сказанного очевидно, что ПЭП растения должно обладать сложной структурой. В работе Безматерных (1982) была обнаружена квазипериодиче- ская структура постоянного электрического поля традесканции. Она была получена в условиях измерения ПЭП симпласта одним электродом, который перемещали вдоль стебля. РЭП каждой исследуемой точки измеряли через 5-10 мин после установления электрода на стебле, т.е. после окончания переходных процессов. Это означает, что в данном случае измеряли не мгновенную картину распределения ПЭП растения. Длительность измерения ПЭП составляла около часа. На рисунке 1 схематично представлена традесканция и распределение РЭП (Δφ) по ее симпласту. Сплошной линией а представлено среднее значение ПЭП неповрежденного растения, пунктирной показаны верхний и нижний пределы его изменения. Ширина распределения ПЭП неповрежденного растения составляет 1,5 мВ и определяется шириной уровня жизнедеятельности растения в переменных внешних условиях: в условиях изменения температуры, уровня освещенности, времени года и т.д. Квазипериодическая структура мгновенного распределения ПЭП, представленная на рис. 1, наблюдается всегда. Как видно из рисунка, РЭП между узлом и корнем на 20-30 мВ меньше, чем между междоузлием и корнем (среднее по 30 растениям). Узел отличается от междоузлия тем, что при помещении его в воду из него растет корень. Таким образом, узел и междоузлие функционально и морфологически различны и различны мембранные потенциалы клеток, образующих их симпласт, т.е. между ними существует РЭП.
Экспериментально установлено, что разность потенциалов между узлом и междоузлием тем больше, чем дальше изучаемое междоузлие расположено от корня. Это означает, что на более молодых междоузлиях большие градиенты потенциалов. Нижние междоузлия имели максимальную РЭП по стеблю, равную 20 мВ, а верхние междоузлия - 36 мВ. Кроме того, на нижних междоузлиях иногда наблюдали два экстремума. Целью данной работы является изучение постоянного электрического поля между узлами и междоузиями традесканции. Приборы и материалы: цифровой вольтметр, традесканция, хлорсеребряные электроды. Методика работы. Один из электродов поместить в первый узел растения, а второй -у основания стебля. Подождать 10 минут до установления равновесия переходных процессов и записать показания вольтметра. Последовательно, через равные промежутки времени, перемещать электрод вдоль стебля шагами на (1/3) длины междоузлия, получить значения электрического потенциала в 7-ми точках, т.е. определить распределение потенциала на двух междоузлиях (между тремя узлами). Данные занести в таблицу.
Построить графическую зависимость электрического потенциала по полученным результатам. Контрольные вопросы: 1.Что такое мембранный потенциал? 2. Что такое вариабельный потенциал и его отличие от потенциала действия? 3. Какова роль мембранного потенциала в симпласте высшего растения? 4. От чего зависит величина мембранного потенциала? 5. На каких участках стебля разность электрических потенциалов будет наибольшей относительно корневой системы растения? 6. Почему наблюдается квазипериодичность в структуре постоянного электрического поля традесканции? Рекомендуемая литература: 1. Электрическая система регуляции процессов жизнедеятельности / Под редакцией Г.Н. Зацепиной. - М.: Изд-во МГУ, 1992. - 160 с. 2. Беспалова С.В., Ульянов А.Н. Электрические явления в биомембранах: Теория и лабораторный практикум.- Донецк: ДонГУ, 1999.- 75 с. 3. Медведев С.С. Электрофизиология растений. Учебное пособие. - СПб. Изд-во: СПб ГУ. 1998. 4. Опритов В.А. Электричество в жизни растений // Соросовский образовательный журнал. - 1996. - №9. - С. 40-46. 5. Опритов В.А. и др. Биоэлектрогенез у высших растений. - М.: Наука, 1991. 5. ИССЛЕДОВАНИЕ ЭЛЕКТРИЧЕСКОЙ АКТИВНОСТИ СЕРДЦА - ЭЛЕКТРОКАРДИОГРАФИЯ Функционирование живых клеток сопровождается возникновением электрических напряжений — биопотенциалов. Клетки, образуя целостный орган, формируют сложную картину его электрической активности. Она определяется как электрической активностью отдельных клеток, так и взаимодействием между ними, устройством самого органа, процессами регуляции в нем и целым рядом других причин. Электрическая активность отражает функциональное состояние клеток, тканей и органов. Регистрация и анализ электрической активности позволяют проводить биофизические и медико-биологические исследования с целью изучения работы органов и проведения клинической диагностики. В процессе жизнедеятельности состояние органа, а следовательно и его электрическая активность, меняются с течением времени. Это вызвано прежде всего распространением волн возбуждения по нервным и мышечным волокнам. В исследовательских целях могут измерять разность потенциалов непосредственно на поверхности или на внутренних структурах изучаемого органа (сердца, мозга и др.). Однако в клинической практике такое «прямое» измерение разности потенциалов на органе трудно осуществимо. Поэтому для оценки функционального состояния органа по его электрической активности используется принцип эквивалентного генератора. Он состоит в том, что изучаемый орган, состоящий из множества клеток, возбуждающихся в различные моменты времени, представляется моделью — единым эквивалентным генератором. Считается, что этот эквивалентный генератор находится внутри организма и создает на поверхности тела электрическое поле, которое изменяется в соответствии с изменением электрической активности изучаемого органа. Исследование работы органов или тканей, основанное на регистрации во времени потенциалов электрического поля на поверхности тела, называется электрографией. Два электрода, приложенные к разным точкам на поверхности тела, регистрируют меняющуюся во времени разность потенциалов. Временная зависимость изменения этой разности потенциалов Δφ (t) называется электрограммой. Наибольшее распространение в медицинской практике в настоящее время получило изучение электрической активности сердца - электрокардиография. Экспериментальные данные показывают, что процесс распространения возбуждения по различным частям сердца сложен. Скорости распространения возбуждения варьируются в сердце по направлению и по величине: так, в стенках предсердий возбуждение распространяется со скоростью 30-80 см/с, в атриовентрикулярном узле - 2-5 см/с, в пучке Гиса - 100-140 см/с. Полное описание электрического состояния сердца, математическое описание распределения мембранных потенциалов по всему объему сердца в каждой клетке и описание изменения этих потенциалов во времени невозможно. Поэтому, в соответствии с принципом эквивалентного генератора, сердце эквивалентным генератором тока, электрическое поле которого близко по свойствам электрическому полю, созданному сердцем. Для расчетов электрического поля, созданного генератором тока в однородной проводящей среде, генератор представляют в виде токового электрического диполя — системы из положительного и отрицательного полюса (истока и стока электрического тока), расположенных на небольшом расстоянии l друг от друга. Важнейший параметр токового диполя — дипольный момент Разность потенциалов двух точек поля точечного электрического диполя, расположенных на одинаковом расстоянии от диполя, пропорциональна проекции дипольного момента на прямую, на которой лежат эти точки. Исследуя изменения разности потенциалов на поверхности человеческого тела, можно судить о проекциях дипольного момента сердца, следовательно, о биопотенциалах сердца. Эта идея положена в основу модели Эйнтховена, голландского ученого, создателя электрокардиографии. Основные постулаты этой модели: 1. 1. Электрическое поле сердца представляется как электрическое поле точечного диполя с дипольным моментом Е, называемым интегральным электрическим вектором сердца (ИЭВС) (складывается из диполей разных частей сердца: 2. ИЭВС находится в однородной проводящей среде, которой являются ткани организма. 3. Интегральный электрический вектор сердца Е меняется по величине и направлению. Его начало неподвижно и находится в атриовентрикулярном узле, а конец описывает сложную пространственную кривую, проекция которой на фронтальную плоскость образует за цикл сердечной деятельности (в норме) три петли Р, QRS и Т. Изменение величины и направления вектора Е за один цикл сокращения сердца объясняется последовательностью распространения волн возбуждения по сердцу: волна начинает распространяться от синусового узла по предсердиям (петля Р), атриовентрикулярному узлу, по ножкам пучка Гиса к верхушке сердца и далее охватывает сократительные структуры к базальным отделам (комплекс QRS). Петле Т соответствует фаза реполяризации кардиомиоцитов. Эйнтховен предложил измерять разности потенциалов между двумя из трех точек, представляющих вершины равностороннего треугольника, в центре которого находится начало ИЭВС. В практике электрокардиографии разности потенциалов измерялись между левой рукой (ЛP) и правой рукой (ПР) -1 отведение, между левой ногой (ЛН) и правой рукой (ПР) - II отведение, между левой ногой (ЛH) и левой рукой (ЛР) - III отведение. Руки и ноги рассматривались как проводники, отводящие потенциалы от вершин треугольника Эйнтховена., располагающихся у мест прикрепления конечностей к туловищу. Электрокардиограмма - это график временной зависимости разности потенциалов в соответствующем отведении, а значит и временной зависимости проекции ИЭВС на линию отведения (рис.1). Регистрируемые разности потенциалов составляют 0,1-5 Мв.
Электрокардиограмма представляет собой сложную кривую с, соответственно петлям, пятью зубцами P,Q,R,S,T и тремя интервалами нулевого потенциала. Электрокардиограмма состоит из 3-х направленных вверх положительных волн - Р, R, Т и двух направленных вниз отрицательных волн - Q и S. Для любого выбранного момента времени направление и модуль интегрального электрического вектора сердца имеют определенную величину, но проекции этого вектора на три отведения различны. Поэтому ЭКГ в I, во II и в III отведениях имеют разные амплитуды и конфигурации одноименных зубцов. Гармонический спектр электрокардиограммы (набор простых синусоидальных колебаний, на которые согласно теореме Фурье, можно разложить сложное колебание) в основном содержит частоты от 1 до 100 Гц, есть и постоянная составляющая. Три отведения не дают полной информации о работе сердца. Поэтому современная кардиология использует 12 стандартных отведений и ряд специальных. ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ Цель работы: Измерение изменения разности потенциалов сердца на поверхности тела и расчет параметров ЭКГ человека. Приборы и материалы: электрокардиограф, электроды, 5-10% раствор NaCl
|

 , где I — ток в диполе равный суммарному току во внешней среде,
, где I — ток в диполе равный суммарному току во внешней среде, 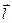 — вектор расстояния между полюсами.
— вектор расстояния между полюсами.




