Г. Н. НикитинаОСНОВНЫЕ ТЕОРЕТИЧЕСКИЕ ПОДХОДЫ К ИЗУЧЕНИЮ ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ РАЗВИВАЮЩЕГОСЯ МОЗГА ЧЕЛОВЕКА [1] Среди важных проблем современной нейробиологии развития особое место занимает проблема функциональной организации мозга человека в раннем периоде жизни. Это связано прежде всего с необходимостью познания становления психической активности ребенка как социального существа. Еще И.М.Сеченов (1958), рассматривая проявления психической деятельности человека от ее истоков, лежащих в младенческом возрасте и проявляющихся в этом периоде индивидуального развития в чувственном возбуждении, и до формирования сложных психических актов, указывал на значение в их реализации функциональной интеграции структур мозга. В исследованиях И. П. Павлова, открывшего феномен условно-рефлекторного поведения, впервые было научно доказано существование зависимости между функциональными проявлениями мозга и психической деятельностью. Однако психическая деятельность человека представляет собой более высокое качество, чем функция структур, лежащих в основе этой деятельности. Этот новый вид деятельности возникает в ходе эволюции на основе всей сложности и многообразия поведения человека, мышления и социального взаимодействия. Развитие его идет в процессе восприятия действительности на уровне внутренних схем — «оперативных констелляций» и в процессе «подстройки» их к конкретным запросам окружающей среды (Кемали, Вакка, 1981). В основе психических процессов лежит активность множества мозговых структур — макро- и микросистем, функционально связанных благодаря тонкой пространственно-временной организации нейрофизиологических процессов. «Высшие психические функции как сложные функциональные системы не могут быть локализованы в узких зонах мозговой коры или в изолированных клеточных группах, а должны охватывать сложные системы совместно работающих зон, каждая из которых вносит свой вклад в осуществление сложных психических процессов и которые могут располагаться в совершенно различных, иногда далеко отстоящих друг от друга участках мозга» (Лурия, 1969). Одним из первых, кто обратил внимание на иерархический принцип локализации психических функций, был И. Н. Филимонов (1940, 1974). Им было сформулировано положение о функциональной многозначности мозговых структур, которые при определенных условиях могут включаться в выполнение тех или иных функций. В пользу этого положения свидетельствовали также работы И. П. Павлова, который, как известно, выделял в коре больших полушарий «ядерные зоны анализаторов» и «рассеянную периферию» со свойственными ей пластическими функциями. В последующем были получены многочисленные физиологические морфологические данные, свидетельствующие о динамичности, изменчивости организации функций мозга. Среди фундаментальных работ в этом направлении следует отметить экспериментальные исследования П.К.Анохина (1968,1971), О. С. Адрианова (1976, 1980), Н. П. Бехтеревой (1971, 1980) и их учеников, показавших, что различные по сложности физиологические функции и поведенческие акты обеспечиваются динамической организацией разнообразных по структуре функциональных систем, включающей аппарат жестких и гибких связей. Пониманию и конкретизации особенностей взаимодействия функциональных процессов, относящихся к разным уровням мозговой организации, несомненно должен способствовать и анализ их морфологических особенностей с позиции учения о макро- и микроансамблевой организации мозга на разных стадиях развития (Адрианов, 1976, 1987). Постепенное и гетерохронное созревание этих систем в ходе индивидуального развития организма создает специфику этого взаимодействия, а, следовательно, и специфику формирующейся интегративной деятельности мозга, обеспечивающей адаптацию в окружающей среде (Орбели, 1961; Анохин, 1971; Волохов, 1980; Хрипкова, Фарбер, 1982, и др.). Для того чтобы понять, хотя бы в основных чертах, каким образом родившийся младенец входит в окружающий его мир и приспосабливается к нему, каковы пути и механизмы этого взаимодействия с постоянно усложняющейся средой, включая социальное окружение, необходимо полностью раскрыть основные теоретические подходы, на которых базируется анализ развития функциональной активности мозга ребенка. Прежде всего это касается проблемы раннего индивидуального опыта и его роли в функциональном созревании мозга, которая остается весьма актуальной и на сегодняшний день. Важность этого направления онтогенетических исследований обусловливается необходимостью кардинального вопроса нейробио-логии — о степени генетической детерминации и воздействий внешней среды в адаптивной деятельности мозга, с одной стороны, и компенсаторных процессах при возникновении нарушений в ходе онтогенеза — с другой. Решение этой задачи тесно связано с экспериментальным обоснованием принципа этапности — одного из основных принципов индивидуального развития функций центральной нервной системы как на системном, так и на клеточном уровне (Богданов, 1864, 1986; Волохов, 1968, 1978; Никитина, 1971, 1979; Аршавский, 1975, и др.). Генетически «запрограммированная» цепь онтогенетических последовательностей, очевидно, является продуктом естественного отбора («стабилизирующего отбора», по Шмальгаузену, 1968), который действовал на гены, оказывающие более или менее прямое влияние на проявление определенных признаков в организме. Однако вследствие недостаточной разработки общей теории индивидуального развития остаются неясными процессы, происходящие в период преобразования наследственной информации в конкретный феномен. Индивидуальное развитие организма всегда зависит от факторов окружающей среды. Поэтому провести определенную грань между признаками и дифференщировками, зависимыми и независимыми от внешних факторов, практически невозможно — онтогенез в этом смысле неразложим (Шмальгаузен, 1968). Концепция периодизации индивидуального развития положена в основу многих исследований по онтогенезу мозга человека и животных (Выготский, 1960; Орбели, 1961; Анохин, 1971; Эльконин, 1971; Волохов, 1975, 1980; Аршавский, 1975, 1982; Фарбер, 1979; Scott, 1975; Любомирский, 1990, и др.). Согласно этой концепции морфофункциональный онтогенез мозга представляется генетически обусловленной последовательностью периодов развития, которая реализуется при взаимодействии организма с условиями среды, соответствующими каждому из этих периодов. Переход от одного периода к последующему, качественно новому периоду развития происходит обычно в сравнительно короткие отрезки времени, которые рассматриваются как переломные или критические периоды онтогенеза. При этом очень существенно, как подчеркивает И. А. Аршавский (1975), что не изменение среды на соответствующих переломных этапах индивидуального развития обусловливает переход от одного возрастного периода к последующему, а изменение самого развивающегося организма, его морфофункциональных систем требует новых условий среды и обусловливает преобразование форм взаимодействия с ними. Именно наличием критических периодов и определяется дискретность непрерывного в своей сути процесса онтогенеза. Возникающие структурно-функциональные перестройки в системах мозга обеспечивают осуществление новых физиологических функций, форм поведения. И от того, как скоро и пластично осуществляются эти адаптивные перестройки, зависит степень физиологической полноценности той или иной системы и организма в целом, а также течение последующих стадий онтогенеза. Это относится, по-видимому, и к пренатальному онтогенезу, в котором закладка структур и функций может рассматриваться не только как преадаптация (предварение будущего), а как форма адаптации к тем специфическим условиям, которые создаются материнским организмом. Значение критических периодов в антенатальном развитии млекопитающих подробно исследовалось П. Г. Светловым и сотрудниками (1978). Результаты этих исследований дали основание считать, что критические периоды в антенатальном развитии — это периоды детерминации, которые отделены от специального функционирования детерминируемых структур более или менее значительным латентным периодом. Для этих периодов характерно повышение чувствительности к тем или иным повреждающим агентам (гипоксия, гипотермия, радиация и др.). У млекопитающих и человека стадии повышенной чувствительности к указанным факторам совпадают с имплантацией плода, органогенезом и плацентацией. Они связаны с особенностями обмена веществ, в частности усилением окислительных процессов, изменением содержания рН-групп и цитоплазматических нуклеотидов и др. («Развивающийся мозг и среда». М., 1975). В последние годы понятие «критические периоды» обосновывается и находит свое приложение в изучении этапности развития физиологических и психологических функций у нормально развивающегося организма человека и животных в пренатальном и постнатальном онтогенезе. Оно широко используется для анализа изменений в организации любой живой системы — процесса роста, развития сенсорных способностей, поведения, изменений в социальной системе (Scott, 1975). При этом разграничение критического периода зависит от сущности процесса и его относительных временных границ. В классическом случае критического периода процесс преобразования системы происходит в течение короткого интервала времени, а затем полностью прекращается, что характерно для многих эмбриональных процессов. Иную временную динамику имеют процессы социального привыкания у высших животных и человека; в этом случае процесс вначале также может протекать быстро, затем он снижается до определенного уровня, но полностью не прекращается. Имеется еще одна возможность динамики критического периода — когда организационные процессы могут происходить периодически с большой скоростью и, следовательно, с многократными ритмическими критическими периодами. Таким является процесс обучения у высших животных. Следовательно, в определении критического периода в онтогенезе той или иной биологической системы ведущее значение имеет критерий скорости (время) развития процесса. Другим важнейшим фактором преобразования системы и перехода ее на новый уровень организации является возникновение повышенной чувствительности к воздействиям среды определенного типа, т. е. избирательной. Действительно, именно в тот момент, когда функция бурно развивается, ей необходим адекватный сенсорный приток, и его отсутствие или дефицит может оказаться фатальным для становления данной функции. Это положение иллюстрируют схема (рис. 1), в основу которой положена общебиологическая концепция Дж. Скотта (1975) о процессах организации системы, а также данные других авторов. Из приведенного рисунка видна тесная взаимосвязь критического и сенситивного периодов в развитии системы. При этом вследствие повышенной избирательной чувствительности системы к средовым воздействиям, возникшей в сенситивном периоде, он может быть одновременно и оптимальным периодом Для направленных воздействий с целью оптимизации или коррекции развивающихся процессов, и периодом уязвимости системы к повреждающим воздействиям. А повышенная чувствительность к повторяющимся сенсорным стимулам может быть начальным проявлением селективности системы, которая должна сформироваться в последующем периоде, требующем более высокого уровня адаптации к среде. Как известно, концепция сенситивных периодов, которая впервые была предложена Дж. Готлиебом (Gottlieb, 1976), использовалась для анализа возможных исходов перцептивного развития в зависимости от взаимодействия генотипа и средового опыта. Показано, что в период развития, предшествующий началу специфического опыта, например до рождения или до открытия глаз (для зрения), тем не менее существует возможность полного или частичного развития фенотипа. Однако специфическая природа и время сенситивного периода не были объединены автором в общую модель перцептивного развития на основе определенных количественных критериев. А
Рис. 1. Схема, иллюстрирующая возможные варианты развития системы в критическом сенситивном периоде в' зависимости от генетических и средовых факторов (составлена по данным J. Scott, 1975). 1 — благоприятный эффект, направленные воздействия; 2 — неблагоприятный эффект, ведущий к задержке и отклонениям в ходе развития системы Это сделано в работах Р. Эйслина (Aslin, 1981), который представил сенситивный период в виде средового фильтра, воспринимающего одни воздействия и блокирующего другие. Исход развития при этом зависит от ряда факторов, таких, как уровень зрелости фенотипа, адекватность стимула и своевременность его воздействия, фильтрующих характеристик сенситивного периода, а также эластичности системы, которая способна нивелировать в определенных пределах депривационные последствия. Предложенная схема была использована для описания возможных онтогенетических исходов восприятия речи младенцами и их связи со способностью дискриминировать различные фонетические контрасты, а также для развития бинокулярного зрения (Aslin & Pisoni, 1980; Aslin, Dumnaris, 1980}. Из представленных на рис. 2 нескольких возможных исходов перцептивного развития в зависимости от уровня созревания фенотипа и различных средовых входов видно, почему средовые влияния способствуют лишь сохранению бинокулярности у кошек (кортикальная бинокулярность полностью представлена у котят к моменту открытия глаз) и, напротив, являются условием для формирования бинокулярного зрения у человека, так как стереопсис начинает проявляться в 3 — 4 месяца после рождения. Рис. 2. Возможные исходы развития перцептивных способностей в зависимости от уровня развития фенотипа, предшествующего началу опыта, и средовых воздействий. А— созревание функций; Б — сохранение на максимальном уровне; В — облегчение; Г — настраивание (коррекция); Д— индукция. По вертикали —уровень развития перцепции от О до максимума, по горизонтали — динамика воздействия средового опыта; 1 — наличие, 2 — отсутствие средового опыта, пунктирная линия — начало опыта (по: R. N. Aslin, 1981)
Модель «фильтра» Р. Эйслина дает возможность, понять основные механизмы, определяющие избирательность, реагирования развивающейся системы при наличии широкого диапазона доступных средовых входов. Предполагается, что существует генетически специфичный диапазон «приемлемых» средовых входов, который включает базовые требования, необходимые для развития каждой конкретной системы. Этот диапазон рассматривается как фильтр, характеристики пропускания которого настроены на определенные виды средовой стимуляции, оказывающие решающее воздействие на процесс фенотипического развития данной функции, происходящий в рамках сенситивного периода. Например, известно, что зрительное предпочтение специфических лиц возникает у младенцев спустя несколько месяцев после рождения (Nelson, 1987). В то же время «низкоуровневый» фенотип, например внимание ребенка к контуру, возникает раньше, чем фенотип предпочтения лица. При депривации контура в раннем постнатальном онтогенезе может пострадать и развитие лицевого предпочтения. Дефицит в этом позднем фенотипе является результатом неспособности низкоуровневого фенотипа изменить характеристики фильтра для фенотипа более высокого уровня (лицевые предпочтения) (Aslin, 1981). Схематическое изображение сенситивного периода, характеризующегося повышенной избирательной чувствительностью к входящим в диапазон «приемлемых» средовых входов воздействиям, представлено на рис. 3.
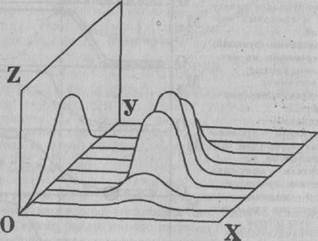
Рис. 3. Схема сенситивного периода, отражающая зависимость от возраста избирательной чувствительности перцептивной системы к воздействиям, входящим в генетически специфичный диапазон «приемлемых» средовых входов. По осям: X — последовательность доступных средовых входов, Y — возраст, Z — чувствительность перцептивной системы (по: R.N.Aslin, 1981) Оценивая критические и сенситивные периоды прежде всего с точки зрения их биологической целесообразности, необходимо подчеркнуть, что они являются узловыми точками развития, дающими импульс коренным преобразованиям в различных звеньях развивающихся систем; за этими процессами следует период более медленных изменений и консолидации системы. В определенных точках развития критический и сенситивный периоды совпадают по времени, что существенно для характера конечного результата. Исходя из сказанного, можно полагать, что происходящие в критические периоды морфофункциональные преобразования в центральной нервной системе носят в основном пластический характер. Но поскольку эти процессы включают и элементы дезорганизации механизмов, свойственных предыдущему возрастному этапу, то они могут оказаться и наиболее уязвимыми при неблагоприятных воздействиях. Поэтому рассмотрение нейрофизиологических, морфологических и психологических данных в разрезе возрастной периодизации представляется важным не только для построения целостной схемы онтогенеза функций мозга, но и для сравнительной объективной оценки уровня развития системной деятельности мозга в норме и при начинающейся патологии нервно-психической сферы ребенка, которая может проявляться в нарушении общего хода, дезинтеграции развития этих функций. Анализируя проблему возрастной периодизации, нельзя не коснуться вопроса о соотношении генетических и средовых факторов в развитии физиологических систем, включая мозговую деятельность. Как уже отмечалось, любое развитие обусловлено реализацией как генетических, так и эпигенетических факторов. Цепь от гена к признаку многоступенчата и состоит из многих сложных и взаимообусловленных процессов. При этом становление одних признаков мало зависит от среды, а других — в значительной степени обусловлено внешними факторами, что прослеживается на различных уровнях организации системы (Богданов и др., 1986). Поэтому разделить влияние генетических и средовых факторов в ряде случаев не представляется возможным, особенно это касается микросреды (Кокина, 1980). Имеется периодизация генной активности, когда чередуется во времени и пространстве то жесткая детерминация, то гибкая, что характерно не только для клеточного уровня, но и для целостной системы. Эти данные подтверждают основной тезис: генетически запрограммированное развитие включает процесс развития, в той или иной степени обусловленного внутренними и внешними связями. Активирующим фактором структурно-функционального созревания специфических свойств центрального аппарата является сама развивающаяся функция (Фарбер, 1969; Богданов, 1978; Барсова, Писарева, 1980; Никитина и др., 1980, и др.). О. В. Богдановым (1978, 1984) на основании обобщения большого экспериментального материала по развитию двигательной функции в эмбриональном и раннем постнатальном периодах онтогенеза человека и животных высказано мнение об афферентно зависимом структурно-функциональном развитии мозга. Им обоснованы некоторые эволюционные закономерности, в том числе роль двигательной афферентации как эндогенного механизма структурно-функциональных преобразований. При этом различные варианты направленных перестроек под влиянием строго учитываемого оптимума афферентных воздействий рассматриваются как первые этапы формирования индивидуального опыта.
|