III.1. Вклад Пн-выбора и фенотипического отбора в приобретение и потерю фенотипически значимых генов.Рассмотрим роли фенотипического отбора и Пн-выбора при утрате и приобретении фенотипически значимых генов и фрагментов ДНК, не влияющих на фенотип.. Под фенотипической значимостью гена подразумевается возможность селекции кодируемого им признака. В случае приобретения нового гена реципиентным геномом положительный результат (внедрение), независимо от фенотипической значимости привносимой информации, будет зависеть от наличия сайта интеграции для конкретного полинуклеотидного фрагмента (нового гена) и наличия ферментов, обеспечивающих этот тип интеграции. То есть в данном случае необходимым и достаточным является Пн-отбор. Фенотипический естественный отбор может служить только дополнительной гарантией стабильности унаследованного гена, пока существуют условия, в которых продукт гена значим для клетки. К подобному заключению относительно градации ролей Пн-отбора и естественного отбора пришли и другие авторы, работающие с растениями [20]. То есть, фенотипический отбор не является необходимым для наследования и сохранения в геноме генетического материала. Подтверждением этого является повсеместное присутствие в клетках самых разнообразных не кодирующих последовательностей. Достаточно сказать, что псевдогены и не кодирующие повторы, хотя и в очень незначительном количестве, обнаружены даже в одном из самых маленьких из известных на настоящий момент геноме Nanoarchaeum equitans — единственного паразита среди архей и единственного представителя нового царства наноархей. При этом геном N. equitans не только очень маленький, но и самый компактный (95% ДНК кодирует белки и стабильные РНК), в отличие от геномов бактериальных паразитов и эндосимбионтов, не претерпел редуктивной эволюции, а был сформирован в теперешнем виде изначально. Авторы, правда, полагают, что N. еquitans является, все-таки, производным какой-то более древней археи. (It has a highly compact genome with few pseudogenes or long regions of noncoding DNA. Consequently, we suggest that this microbe is a derived, but genomically stable parasite that diverged anciently from the archaeal lineage). [109]. (Здесь необходимо отметить, что только что была опубликована на данный момент сенсационная статья, в которой показано существование бактериального организма, геном которого значительно меньше, чем геномы всех (трёх) ранее известных «рекордсменов». Это эндосимбионт, живужий в цитрусовых псилидах — насекомых, питающихся соком растений. Он назван Carsonella ruddii [71]. Геном этой бактерии состоит только из примерно 160 т.н.п. и содержит только 182 белок-кодирующих генов. Это очень близко к рассчитанному на компьютере минимальному размеру генома — 113 т.н.п. Уместно напомнить, что геном человека состоит из 3 миллиардов н.п.) Геномы, претерпевшие редуктивную эволюцию в значительном объёме, тоже содержат псевдогены. У B. aphidicola str. Bp из Baizongia pistaciae их 10, а это значит, что ненужные гены не были утрачены как не поддерживаемые отбором, а в течение миллионов лет сохранялись в предельно редуцированном геноме. Наиболее яркие примеры наследования генов, которое трудно объяснить влиянием фенотипического отбора, то есть предоставлением селективного преимущества бактериям, унаследовавшим новый ген, предоставляет горизонтальный перенос. 1). Бактерии B. quintana, содержат горизонтально перенесенный ген yopP, кодирующий антифагоцитарный белок-эффектор иерсиний, и ген секреторного белка для гемолитического токсина азотфиксирующей почвенной бактерии Sinorhizobium meliloti. Ни тот, ни другой ген не могли быть селекционированы фенотипическим отбором, поскольку являются частями сложных генетических систем и сами по себе никакой функции выполнять не могут. Однако оба этих гена сцеплены с генами тРНК и, следовательно, внедрились в сайт интеграции тРНК [9]. 2) Ген пролил-тРНК синтетазы M. leprae отличается от других генов тРНК синтетаз этой бактерии. Он происходит не от общего с M.tuberculosis предшественника, а скорее всего был перенесен из генома Borrelia burgdoferi [34]. 3). Одноклеточные простейшие, — патогенные для человека парабазилиды Trichomonas vaginalis — получили гены прoлил-тРНК-синтетазы и аланил-тРНК-синтетазы от представителя иного царства (Nanoarchaeota) — N. equitans [14]. 4). Ещё один случай перемещения генетического материала между царствами: фрагмент генома бактерии Wolbachia встроился в X-хромосому жука Callosobruchus chinensis [72]. Указанные случаи, однако, легко объясняются на основе концепции Пн-отбора. При потере гена складывается другая ситуация. Для удаления генетического материала из ДНК также необходим Пн-выбор. В данном случае он выражается в наличии фланкирующих удаляемый ген полинуклеотидных последовательностей, благоприятствующих исключению. Наиболее изученные случаи — это наличие повторяющихся последовательностей, типа IS-элементов, на которых могут оперировать те или иные ферменты рекомбинации. Если гены сохраняются в геноме, это не означает автоматически, что они необходимы для клетки в данных условиях. Это означает, что в геноме нет условий для их утраты. Так, показано, что потеря генов при редукции геномов происходит в два этапа. Сначала ген инактивируется за счёт мутации, а потом уже инактивированный ген (псевдоген) делетируется [10, 97, 48]. Период полураспада псевдогенов у Buchnera aphidicola составляет 24 миллиона лет. Полная элиминация псевдогена достигается за 40 — 60 миллионов лет [48]. То есть, ненужная ДНК задерживается в геномах на десятки миллионов лет. Среди патогенных бактерий, геном которых в наибольшей степени «пострадал» от редукции, выделяется M.leprae (Табл. 1). Несмотря на утрату значительной части, редуцированный геном M.leprae сохранил 1 115 псевдогенов — намного больше, чем в геноме любого другого возбудителя и достаточно большое количество некодирующих повторов (Табл. 1, Рис. 7). Как в геноме Y.pestis, находящейся, видимо, в начальной стадии редуктивной эволюции, так и в геноме M.leprae, ушедшей по пути редукции генома дальше других возбудителей инфекций, IS-элементов намного больше, чем у их «не редуцированных» ближайших родственников (см. выше). Сохранение и «размножение» в геномах Y.pestis, B.pertussis, B.mallei, M.leprae, претерпевающих редукцию геномов, фенотипически не значимой ДНК (повторов различного типа, IS-элементов) также свидетельствует против утверждения, что «ненужный» генетический материал, не поддерживаемый отбором, утрачивается.
Рис. 7. Количество псевдогенов в геномах различных бактерий.
Экспериментальных доказательств того, что удаление из генома нейтральной генетической информации сообщает клеткам селективное преимущество нет или они неизвестны автору. Однако существуют данные о том, что в результате потери генетического материала селективные преимущества не возникают [98]. Если условия Пн-выбора позволяют потерю ДНК за счёт делеции, наследование такой делеции в потомстве будет строго зависеть от того, насколько необходим утраченный признак для клетки в данных условиях существования. То есть, вероятность потери фенотипически не значимых участков генома будет определяться только Пн-выбором. В случае благоприятного для делеции полинуклеотидного окружения данного фенотипически не значимого сегмента ДНК, такой сегмент будет с высокой вероятностью делетирован. Если рассматриваемый сегмент ДНК содержит фенотипически значимую информацию, то гены, кодирующие необходимые на данный момент и в данных условиях признаки, сохранятся в потомстве. Это не значит, что делеций этих генов не происходит. Просто выживают бактерии, не претерпевшие делеций. Примером нестабильности, определяемой контекстом, может служить поведение одного из участков ДНК Y. pestis.
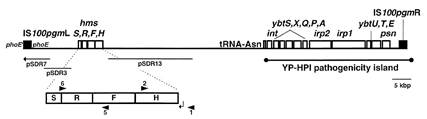
Рис. 8. Pgm локус (102 тнп) Y. pestis KIM10, ограниченный прямыми повторами элемента IS 100.
В хромосоме иерсиний есть локус рgm (Рис. 8). Если все гены этого локуса полноценны, имеет место пигментация бактерий (Pgm+ -фенотип). Однако этот фенотип нестабилен и с большой частотой утрачивается [28]. Оказалось, что весь сегмент ДНК, размером 102 тнп, делетируется за счёт рекомбинации между прямыми повторами ограничивающих его элементов IS 100 [43]. Частота таких делеций в recA+ штаммах достигает значения 10-3 [50]. То есть, такие делеции происходят всё время в каждой тысячной клетке популяции. В отличие от ситуации с псевдогенами B.aphidicola, удаление из геномной ДНК одного из её сегментов не требует не то, что десятков миллионов лет, а происходит постоянно. Следовательно, миллионолетняя задержка удаления псевдогенов имеет место тогда, когда для делеций нет условий. В случае с локусом рgm такие условия есть, — это фланкирующие элементы IS 100. То есть, в случае потери фенотипически значимых генов для осуществления делеции важны условия Пн-выбора, а для сохранения популяции в условиях вероятности делеции, — фенотипический (естественный) отбор. Сделанные выводы суммированы в Табл. 5. Таблица 5. Зависимость приобретения и потери фенотипически значимых генов и нейтральной генетической информации от полинуклеотидного отбора и фенотипического (естественного) отбора.
Из сказанного следует, что генетический материал наследуется извне или удаляется из генома, если для этого есть возможность, определяемая структурой самого генома. Унаследованный ген стабилен, если геном не предоставляет ему условий для делеции. Роль фенотипического (естественного) отбора в этой системе равна нулю. Это означает, что первичные этапы любых изменений геномов и как следствие — выбор пути эволюции геномов (компактизация или накопление) определяются особенностями структуры самих геномов. Структура геномов формируется благодаря предшествующему Пн-выбору. Фенотипический (естественный) отбор оперирует на следующем этапе и его объектом может быть часть тех изменений генома, которые были разрешены его структурой.
|