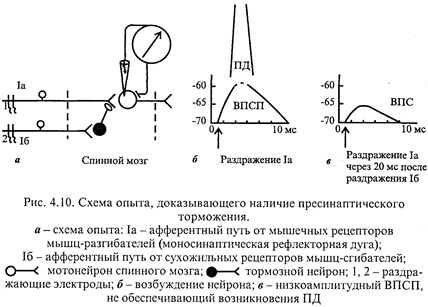
Пресинаптическое торможение. Роль различных видов торможенияА. Открытие. Пресинаптическое торможение первоначально выявлено в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов. Так, известно, что раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных α-мотонейронов (α-мотонейронов этой же мышцы). Однако опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемых α-мотонейронов (рис. 4.10). Интересным в данном случае было то, что мембранный потенциал и возбудимость исследуемых α -мотонейронов не изменялись либо регистрировался низкоамплитудный ВПСП, недостаточный для возникновения ПД. В опыте исследовались мотонейроны в составе моносинаптической рефлекторной дуги, поэтому было очевидно: они не возбуждаются вследствие процессов, происходящих в пресинаптическом окончании, что определяет название этого вида торможения.
Б. Механизм пресинаптического торможения. Электрофизиологическое изучение процессов на уровне пресинаптических окончаний в описанном опыте показало, что здесь регистрируется выраженная и продолжительная деполяризация, ведущая к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения, следовательно, поступающие импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуде, не обеспечивают выделение медиатора в синаптическую щель в достаточном количестве: нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях аксона-мишени (рис. 4.10). Торможение (деполяризация) после одного афферентного залпа продолжается 300-400 мс, медиатором является γ-аминомасляная кислота (ГАМК). Формирование деполяризации пресинаптического окончания на ионном уровне изучено недостаточно. Полагают, что деполяризация является следствием повышения проницаемости для ионов Сl -, в результате чего они выходят из клетки. По-видимому, в составе мембран пресинаптических терминалей имеется хлорный насос, обеспечивающий избыточное поступление ионов Сl - внутрь клетки вопреки электрическому градиенту. Под действием ГАМК тормозных нейронов и последующего повышения проницаемости мембраны для ионов Сl - последние начинают выходить наружу согласно электрическому градиенту. Это приводит к деполяризации пресинаптических терминалей и ухудшению их способности проводить импульсы. Полагают также, что деполяризация пресинаптических терминалей может возникнуть при накоплении К+ в межклеточной жидкости в результате повышенной активности нервных окончаний и соседних нервных клеток. В этом случае также ухудшается проводимость пресинаптических терминалей вследствие устойчивого снижения мембранного потенциала в связи с уменьшением концентрационного градиента дляК+ и уменьшением выхода его из клетки В. Разновидности пресинаптического торможения. Обычно выделяют параллельное и латеральное пресинаптическое торможение. Структурная организация этого торможения внешне похожа на такие же варианты постсинаптического торможения. Однако напомним: механизм пресинаптического торможения отличается принципиально - возбуждение задерживается не на теле нейрона, а на подступах к нему. В случае постсинаптического торможения заторможен нейрон - снижена возбудимость тела нейрона (обычно вследствие его гиперполяризации). В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рис. 4.9. Тем не менее все варианты пре- и постсинаптического торможений можно объединить в две группы: 1) когда блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение) и 2) когда блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов или афферентных импульсов от проприорецепторов с включением тормозных клеток (латеральное и прямое торможение). Активация тормозных клеток может облегчить распространение возбуждения, поскольку тормозные клетки сами могут быть заторможены другими тормозными нейронами с помощью механизмов пре- и постсинаптического торможения (торможение торможения). Г. Локализация в ЦНС пре- и постсинаптического торможения. Оба механизма торможения широко представлены в различных отделах ЦНС: в частности, глицин - медиатор постсинаптического торможения, кроме клеток Реншоу, имеется в ретикулярной формации ствола мозга; ГАМК - в спинном мозге, мозжечке, коре большого мозга. Оба вида торможения могут быть заблокированы: постсинаптическое - стрихнином; пресинаптическое - бикукулином. Пост- и пресинаптическое торможение блокируется также столбнячным токсином, который нарушает высвобождение тормозных медиаторов. Иногда выделяют в качестве разновидности центрального торможения торможение вслед за возбуждением. С точки зрения имеющихся фактов, особым механизмом торможения его считать нельзя, поскольку оно является результатом следовой гиперполяризации нейронов. Если выделять этот вид торможения, то его целесообразно называть «следовое торможение» как результат следовой гиперполяризации нейрона. Пессимальное торможение (пессимум Введенского), наблюдаемое в эксперименте на нервно-мышечном препарате, в ЦНС в физиологических условиях, по-видимому, не встречается. Д. Роль торможения. 1. Оба известных вида торможения со всеми их разновидностями выполняют охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов и прекращению деятельности ЦНС. 2. Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко выражена эта роль у пресинаптического торможения. Оно более точно регулирует процесс возбуждения, поскольку этим торможением могут быть заблокированы отдельные нервные волокна. К одному возбуждающему нейрону могут подходить сотни и тысячи импульсов по разным терминалям. Вместе с тем число дошедших до нейрона импульсов определяется пресинаптическим торможением. Торможение параллельных (латеральных) путей обеспечивает выделение существенных сигналов из фона. Блокада торможения ведет к широкой иррадиации возбуждения и судорогам (например, при выключении пресинаптического торможения бикукулином). 3. Торможение является важным фактором обеспечения координационной деятельности ЦНС.
|