
ПРОЯВЛЕНИЕ ВТОРИЧНОЙ СТРУКТУРЫ БЕЛКОВ В УФ СПЕКТРАХКогда пептидные цепи формируются в упорядоченные структуры (например, спираль) возникает вторичная структура белка. Появление таких структур можно выявить методом УФ спектроскопии. Так, например, на рис. 2.3.1 показано влияние рН и температуры на вид спектра поли-L-лизина. При повышение величины рН (при постоянной температуре) количество положительных зарядов на боковых цепях лизина уменьшается. Это способствует укладке полипептидной цепи в более упорядоченную структуру (спираль). В УФ спектре такая перестройка вторичной структуры (от статистического клубка к спирали) проявляется гипохромным эффектом. Повышение температуры при постоянном рН преобразует спиральную структуру полипептида в β-структуру складчатого типа. Эта перестройка проявляется гиперхромным эффектом. Белки обычно содержат одновременно несколько элементов вторичной структуры в различных положениях пептидной цепи. Процесс денатурации уничтожает значительную часть этих вторичных структур, и как следствие мы имеем изменения в спектрах поглощения. Особенности поглощения, связанные с α-спиралью, β-складчатой структурой или произвольным скручиванием достаточно отчетливы, чтобы можно было использовать их для диагностических целей в случае природного белка, однако здесь более информативными могут быть спектры кругового дихроизма.
Интенсивное поглощение всех нуклеиновых кислот в ближней УФ-области почти целиком обязано пуриновым и пиримидиновым основаниям. Сахарофосфатный остов РНК и ДНК дает незначительный вклад в спектр поглощения при длинах волн, превышающих 200 нм. Спектры рибонуклеотидов весьма близки к ним, за исключением спектра уридина (U), у которого λmах = 260, а не 268 нм. Однако электронная структура пуринов и пиримидинов гораздо сложнее, чем белковых хромофорных групп. Основания имеют очень низкую симметрию и много несвязывающих электронов. Сопоставление со спектрами ряда более простых молекул показывает, что у всех оснований в области от 200 до 300 нм должно проявляться несколько разных n → π*- и π → π*-переходов. Кажущиеся простыми полосы поглощения аденина (A) и урацила (U) с λmax = 260 нм, имеющие вид гауссовых кривых, в действительности порождаются более чем одним переходом. G и С имеют по две полосы. На самом деле они, вероятно, обладают большим числом неразрешенных полос. Свободное основание, нуклеозид (основание, присоединенное к сахару), нуклеотид (основание, присоединенное к фосфату сахара) и денатурированный полинуклеотид все они имеют очень похожие спектры поглощения в этой области (см. табл. 2.3.2). Таблица 2.3.2
Спектры всех четырех нуклеозидов чувствительны к рН. Протонирование С и G приводит к значительному сдвигу поглощения в сторону больших длин волн (красное смещение). Депротонирование U или Т при щелочных рН также приводит к существенному красному смещению максимума поглощения. Протонирование А сопровождается гораздо меньшими спектральными изменениями. Все эти эффекты весьма полезны для определения степени ионизации отдельных составляющих нуклеиновых кислот, однако в полимерах они становятся более сложными из-за сильного электронного взаимодействия между основаниями. Присутствие фосфатных групп, по-видимому, не сказывается сколько-нибудь заметно на молярной экстинкции различных составляющих нуклеиновых кислот. Энергия длинноволновых, достаточно интенсивных электронных переходов пяти наиболее распространенных оснований почти одинакова. Это служит серьезным препятствием для детального анализа электронных спектров ДНК и РНК. Полосы поглощения отдельных хромофоров типичной нуклеиновой кислоты сливаются и дают простую на вид полосу с λmax = 260 нм (см. рис. 2.3.8).
Рис.2.3.8 Спектр поглощения ДНК как функция температуры. После расщепления ДНК ферментами в растворе должны присутствовать только мононуклеотиды и, возможно, небольшое количество коротких олигомеров. Спектр, измеренный при высокой температуре, очень близок к спектру препарата, обработанного ферментами. I - нативная ДНК, 250С; II - препарат, обработанный ферментами; III - ДНК Rcoli, 82oС. [D.Voet, W.B.Gratzer, R.A.Cox, P.Doty, Biopolymers, 1, 193 (1963).] Свойства отдельных оснований оказываются весьма близкими, так что даже в отсутствие достаточно сильного взаимодействия между основаниями трудно представить полосу поглощения с λmax = 260 нм для нуклеиновой кислоты в виде суммы четырех составляющих с определенными весами. Средний коэффициент молярной экстинкции при 260 нм в расчете на моль нуклеозидов равен примерно 1,0·104. В общем, поленуклеотиды и нуклеиновые кислоты поглощают меньше в расчете на нуклеотид, чем сумма составляющих их нуклеотидов. Таким образом, природная двухспиральная ДНК поглощает меньше на нуклеотид, чем денатурированная («расплавленная») нить ДНК, т.е. и здесь, как и в белках, наблюдается гипохромизм.
Рис.2.3.9 Гиперхромный эффект при денатурации нуклеиновых кислот.
Рис.2.3.10 Определение температуры плавления вторичной структуры ДНК Гипохромность полинуклеотидов или нуклеиновых кислот относительно мономерных нуклеотидов является следствием взаимодействий между соседними основаниями при их упорядоченном расположении в спиральной цепи полимера. Происхождение гипохромизма по своей природе электромагнитно. Оно включает взаимодействия между переходными дипольными моментами индивидуальных оснований и их соседей. Таким образом, оно зависит не только от внутренних переходных моментов каждого основания, которые для химически различных оснований отличаются по величине и направлению, но также от относительной ориентации взаимодействующих оснований. Упорядоченные основания поглощает слабее в расчете на нуклеотид, чем неупорядоченные. Стопка из двух оснований имеет немного меньшую оптическую плотность, чем удвоенная оптическая плотность одного основания; и так далее. Таким образом, пары контактирующих оснований в двойной спирали поглощают меньше, чем частично контактирующие основания в одиночной спирали, которые поглощают слабее, чем мононуклеотиды.
|
 Полосы поглощения нуклеотидов
Полосы поглощения нуклеотидов

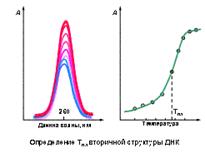 Гипо- и гиперхромизм полинуклеотидов или нуклеиновых кислот относительно нуклеотидов в первую очередь является следствием взаимодействий между соседними основаниями при их упорядоченном размещении в спиральном полимере. Изменения при денатурации в одиночных спиралях или при гидролизе мононуклеотидов легко измеряются (обычно они составляют 30 - 40%) и часто используются для наблюдения за кинетикой или термодинамикой процесса денатурации НК или определения температуры плавления вторичной структуры (см. рис. 2.3.10).
Гипо- и гиперхромизм полинуклеотидов или нуклеиновых кислот относительно нуклеотидов в первую очередь является следствием взаимодействий между соседними основаниями при их упорядоченном размещении в спиральном полимере. Изменения при денатурации в одиночных спиралях или при гидролизе мононуклеотидов легко измеряются (обычно они составляют 30 - 40%) и часто используются для наблюдения за кинетикой или термодинамикой процесса денатурации НК или определения температуры плавления вторичной структуры (см. рис. 2.3.10).


