
Эквивалентті схемаларБиологиялық объектілерден электр тогы өткенде тірі клетканың кедергісін Сиымдылық пен Омдық кедергілердің қосындысы түрінде қарастыру келісілген. Тұрақты токқа клетка мен ткандердің кедергілерінің өте үлкен болуын – конденсатордың тұрақты токты өткізбеуінен деп түсіндіріледі. Ал айнымалы токты қолданған кездегі алынған нәтижелерді түсіндіру үшін, омдық кедергі мен сиымдылық кедергісін есепке алуымыз керек. Омдық кедергі денеден өтетін токтың жиілігіне байланысты болмайды, ал сиымдылық кедергісі жилік көбейген сайын кемиді, ал бұл барлық клетка мен такандердің электр өткізгіштігін үлкейтеді. Сонымен клетка мен тканның электр өткізгіштік дисперция құбылысы ток жилігі көбейген сайын сиымдылық кедергісінің кему нәтижесімен түсіндіріледі. Өте үлкен жиліктерде клетка мен ткандердің өткізгіштігінің өсуі онша көп болмай – тұрақты болып қалады, ал 106 – 108Гц жилікте максимум өткізгіштік пайда болады, себебі бұл кезде сиымдылық кедергісі практика жүзінде 0 – ге тен болады. Қалыпты жағдайда биологиялық системаның осы жиліктегі токқа кедергісі клеткадағы еркін электролиттердің концентрация мәнімен анықталады. Биологиялық системада сиымдылық элементтерінің болуы фаза ығысуының пайда болу нәтижелерімен түсіндіріледі. Егер синусойдалық айнымалы ток тізбегінде тек қана Омдық кедергі болса онда фаза ығысуы болмайды. Бұл жағдайда ток күшінің жарты период ішіндегі өзгеру қисығы осы кездегі кернеудің өзгеру қисығымен дәл келеді, яғни ығысу фазасының бұрышы 0 – ге тең болады. Сиымдылық кедергісінен айнымалы ток өткен кезде ток күші максимал мәнге ие болады да айнымалы ә.қ.к -ің шамасы өзгеріп тік төмендеп немесе тік жоғарылап кетеді. Кернеудің өзгеруі бас нүкте арқылы (0) өткенде ток 1 /4 периодқа кернеуді басып озып, яғни фазасының ығысу бұрышы 900 -қа тең болады. Сиымдылық және Омдық кедергілері бар системада фазалық ығысу бұрышы олардың қатынасымен анықталады.
4.4.-сурет. Конденсатор арқылы айнымалы ток өткенде тоқ күші (J) мен электр өрісі кернеулігінің (Е) өзгеру қисығы. Биологиялық системалар үшін-ығысу фазасының бұрышы үлкен мәндерге ие болады. Ал бұл биологиялық объектілерде сиымдылық кедергісінің үлесі зор болатындығын білдіреді. Мысалы: 103 Гц жиіліктегі кейбір биологиялық объектілердегі алынған фазалық ығысу бұрышы мынандай. 1. Құрбақа нервісі - 640 2. Қоян бұлшық еті - 650 3. Құрбақа терісі - 550 4. Адам терісі - 550 5. Ламинария - 780 Бұл келтірілген мәліметтер - ток жилігінің кең көлемде өзгеруіне қарамай – тұрақты болып қала береді. Себебі биологиялық объектілер – сиымдылық – Омдық жүйеге жатады, ал олардағы сиымдылық - негізінен – статикалық емес – Поляризациялық болып келеді. Статикалық сиымдылығы бар жүйеде жилік көбейген сайын – олардың өткізгіштігі өседі, сонымен қатар фазаның ығысу бұрышы да өседі. Поляризациялық сиымдылығы бар жүйеде – сиымдылық арқылы және Омдық кедергі арқылы ағып өтіп жатқан ток шамаларының қатынасы жиліктің белгілі бір шегінде тұрақты болып қалады. Тірі клеткалардың толық кедергісі – импеданс деп аталады. Электротехника мен физикада – Импеданс деп – активті және рактивті кедергіден тұратын эффективті кедергіні айтады. Ал реактивті кедергі – сиымдылық кедергісі мен индуктивті кедергіден тұрады. Биологиялық объектілерде өзіндік индукция болмайтындықтан импеданс деп тірі клетканың Омдық және сиымдылық кедергілерінің геометриялық қосындысын айтамыз. Тірі клеткалардың электр тогын өткізуін сипаттау үшін, эквиваленттік схемаға жүгінеді, яғни Омдық және сиымдылық кедергілерінің комбинациялары арқылы клетканың электрлік параметрлерін моделдеуге тырысады. Жай эквиваленті схемалардың қарастырайық: 1)
Омдық кедергі мен сиымдылық кедергілері тізбектей қосылған жағдайда олардан өтетін токтың шамалары бір – біріне тең болады. JR = JC. Сол кедергілерде болатын кернеудің шамалары мынаған тең болады.
UR =JRR, UC= JCXC (2) мұндағы; Жалпы кедергілерге түсірілген кернеу – векторлық түрде мынаған тең
Скалар түрінде U2 = J2R R2 + J2c X2c (5) бұдан Сонда толық кедергі – Импеданс Ζ - мынаған тең
Ығысу фазасының тангенс бұрышы мына теңдеумен анықталады.
2) Кедергілер параллель жалғанған жағдайда кедергілерге түсірілетін кернеудің шамалары бір – біріне тең бірдей болады. U = UR =UC
Ал жалпы ток 2- ге жіктеледі.
Сонда
Яғни Импеданс мынаған тең болады:
Жалпы түрде Ығысу фазасының бұрыштық тангенсі мына теңдеумен анықталады. Келтірілген осы жай 2- схеманы толығымен тірі клеткаларға қолдануға болмайды, себебі бұл 2- схемадағы қолданылатын жиліктер диапазоны тірі клеткадағы кедергілер өлшенетін жилікке сай келмейді. Төменгі жилікте Омдық және сиымдылық кедергілерді тізбектей қосу схемасында импеданстың тәжірибелік мәні мен есептеулік мәндерінің бір –бірінен ауытқулары көп болады. Параллель қосу схемасында жоғары жиліктерде системаның Импедансы 0 – ге ұмтылады. ал тірі биологиялық жүйелердің импеданстық шамасы тек белгілі бір шамаға дейін ғана кемиді. Тірі клетканы сипаттау кедергілердің жай жалғануы арқылы схемаданда көрі күрделі схемамен жүреді. Мысалы: Фрикс пен Морзе қызыл қан түйіршіктерінің кедергілерін өлшей келе оларды сипаттайтын теңдеуді тапқан ол теңдеулерге мына эквивалентті схема сәйкес келген (4.5-сурет-І).
4.5.-сурет. Эквиваленттік схемалар І- Фрикс пен Морзе схемасы ІІ- Петров схемасы ІІІ-Шванн схемасы Схемадағы:
Ri – клетка құрылымының кедергісі.
С – мембрананың сиымдылығы. В.А. Петров (1939) - Нерв арқылы баспалдақты кернеудің өтуін зерттеген – ол электр разрядына ұқсас процесстің жүретінін дәлелдеген. Ол осы процесті сипаттайтын (4.5-сурет-ІІ) схеманы ұсынған. Шванн – бұл процестерді сипаттау үшін күрделі (4.5-сурет-ІІІ)-схеманы ұсынған. Бұл схемалар белгілі бір жағдайда – тірі клеткалардың токты қалай өткізетіндігінің моделі ролін атқарған бірақ олардың біреуі де күрделі биологиялық системалардың өту заңдылықтарын дәл келтіре алмайды. Көптеген зертеулермен дәлелденгендей – тірі клетка мен ткандер үшін Импеданстың өзгеруін айнымалы токтың әр түрлі жйліктерінде анықтауға болады. Бұл кезде реактивті және активті кедергілер өзгереді. Практикада – көбнесе – тірі жүйелердің электрөткізгіштігін анықтауда негізінен компенсацияланатын жағына Омдық кедергі мен Сиымдылық кедергісін параллель жалғанған көпірлік схема қолданылады. Мұндай схема – тірі клеткаға ең 1- ші жақындатылған жағдайды ғана моделдеу үшін қолданылады, бірақ оны қолдану арқылы жақсы нәтижелер алуға болады. Электротехника мен физикадан белгілі – Реактивті және активті кедергілерді параллель жалғаған кезде, ол айнымалы ток жилігіне тәуелді функция болып табылады да мына теңдеу бойынша анықталады.
Сонда импеданс
Импеданстық жиліктік байланыс – тәуелділігі көбнесе дөңгелек – векторлық диаграмма түрінде сипаталады (өрнектеледі). Ол диаграмма мына түрде салынады, абсцисса осіне биологиялық объектінің активті кедергісі - Rω, ал ордината осіне реактивті кедергісінің сол бірдей жилікте табылған шамасы - Х ω салынады.(4.6-сурет) Ең төменгі жилікке сай келетін Активті кедергінің шамасы – координатаның бастапқы осінен алыста орналасады, ал жоғары жилік кедергінің шамасы – координатаның бас осіне жақын орналасқан. Реактивті кедергінің шамасы бұл 2 - шеткі жағдайдада минимал мәнге ие болады. Реактивті кедергінің максимал мәні орта жилік облысында жатады. Себебі аз төменгі жиліктерде – токтың ең көп мөлшері, Омдық кедергі арқылы өтеді, ал жиліктің өсуіне қарай - сиымдылықтан өтетін ток шамасының үлесі де өседі, ал бұл бүкіл системаның электр өткізгіштігінің өсуіне әкеп соғады. Жоғары жилікте – поляризацияланған сиымдылықтың мәні О – ге ұмтылады.
4.6.-сурет. Ұлпаның Импеданс – годографы
Әр бір жұп пар Rω мен Хω - ның мәндері үшін импеданс – векторларды қосу ережесімен анықталады.
Ол графиктік түрде – координатаның бас нүктесінен шығатын түзу кесіндімен анықталады жиліктің өзгеруіне байланысты вектордың бағыты мен ұзындығы өзгереді де аяғы доға мен бітеді. Поляризация коэффициенті сияқты дөңгелек диаграмманы тірі жүйенің өмірсүргіш қаблеттілігін анықтау үшін қолдануға болады. Ал тірі жүйенің физиологиялық күйлерін сипаттау үшін жалпы кедергінің жилікке байланысты графигін қолдану өте қолайлы болады. Бұл жағдайда абсцисса осіне ток жилігінің логарафимі, ал ордината осіне – жалпы кедергінің мәні салынады. Сонымен қатар тірі жүйенің өмірсүргіштік қаблеттілігін сипаттау үшін жилікпен байланысты өзгеретін фазалық ығысу бұрыштың тангенсін алуға болады.
Сонымен қорыта келгенде жануарлар мен адамдар организмі тұрақты токты өз бойынан өткізе алады. Тірі организмде Индуктивтік катушкаға барабар болатын жүйе жоқ, сондықтан организм үшін Индуктивтік кедергі 0 – ге тең. Олай болса тірі организмнің толық кедергісі немесе Импедансы актив кедергі (R) мен сиымдылық кедергісінен (Хс) тұрады. Организмнің импедансы сол организмнің физиологиялық қасиеттеріне тікелей байланысты. Жүрек соғуының өзгеруіне байланысты организмге баратын - таралатын қанның мөлшері де өзгеріп отырады, осыған байланысты организмнің импедансы да өзгереді. Яғни импеданстың өзгеруіне қарап жүрек жұмысын тексеруге болады. Осы әдісті реография деп атайды. Онымен мидың реограммасын (реоэнцефалограмма), жүректің реограммасын (реокардиограмма)т.б. жазып алуға болады.
|
 (1)
(1)



 (3) сиымдылық кедергісі.
(3) сиымдылық кедергісі. (4)
(4) (6)
(6) (7)
(7) (8)
(8)
 мұндағы
мұндағы ал
ал 
 (9)
(9) (10) бұдан
(10) бұдан  (11)
(11)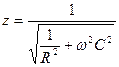 (12)
(12)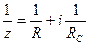
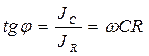 (13)
(13)
 R – клеткааралық сұйықтың кедергісі.
R – клеткааралық сұйықтың кедергісі.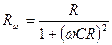 (14) Rω – активті кедергі құраушысы.
(14) Rω – активті кедергі құраушысы. (15) Xω - реактивті кедергі құраушысы.
(15) Xω - реактивті кедергі құраушысы. (16)
(16)

 (17)
(17)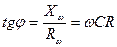 (18)
(18)


