
Кислород и дыхание растений
Экологическое значение кислорода воздуха: – является в клетке главным окислителем, поэтому жизненно необходим для существования эукариотических организмов и многих прокариотов. Их клетки получают основную энергию в процессе кислородного дыхания; – служит конечным акцептором водорода при дыхании; – однако роль свободного кислорода в организмах двойственна. Свободный, атомарный кислород и озон фактически являются клеточными ядами. В атмосфере чистого кислорода дыхание растений снижается, а при длительном его действии растение погибает. Это связано с усилением свободнорадикальных реакций, повреждением мембран вследствие окисления их липидов и в конечном итоге с нарушением многих процессов обмена веществ. Кислород и дыхание растений Дыхание относится к процессам диссимиляции, в которых происходит распад органических субстратов организма. При этом от них отщепляется водород и освобождается некоторое количество энергии, максимальный выход которой связан с переносом водорода на конечный акцептор, который получает его через дыхательную цепь в митохондриях или глиоксисомах. Этот процесс связан с цепью переноса электронов. В дыхательной цепи благодаря фосфорилированию образуется универсальная энергетическая «валюта» клеток – АТФ. Еще высокоэнергетические соединения образуются при фосфорилировании на уровне субстратов. Так как окислительные превращения субстратов при дыхании включают не только аэробные, но и анаэробные процессы, снижение парциального давления кислорода с 21 до 9% незначительно меняет интенсивность дыхания растений. При 5%-м содержании О2 у молодых растений начинает несколько снижаться потребление его и в меньшей степени изменяется выделение СО2. Это связано с тем, что один из важнейших ферментов аэробного дыхания – цитохромоксидаза – обладает высоким сродством к кислороду и может функционировать при его низких парциальных давлениях. Это важный адаптивный элемент надежности энергетики растения, который связан с тем, что газовый состав тканей может отличаться от атмосферного. Так, в листовой паренхиме сахарной свеклы (Beta vulgaris) содержание О2 меняется в течение суток от 7,1 до 17,4%, СО2, – от 0,9 до 5,1%; в зрелых плодах яблони (Malus) содержится 7,5% СО2 и 13,9% О2, у лимона (Citrus) - 8,5% СО2 и 11,5% О2. Поступление кислорода в растение из воздуха и воды Кислород содержится в воздухе в количестве, достаточном для дыхания наземных растений или их наземных частей, и никогда не является для них лимитирующим фактором (исключение – массивные органы с плохой газопроводимостью, например толстые стволы и крупные плоды). Газы попадают в тело растения через устьица, потом растворяются в жидкой фазе клеточных стенок и оттуда проникают в клетку. Этот процесс требует сохранения влажности контактирующих с атмосферным воздухом оболочек и мембран. Поступление кислорода изменяется при механических воздействиях на растение. Например, при этом листья кратковременно (от нескольких минут до 1 ч) усиливают поглощение кислорода, причем надавливание влияет мало, изгибание – сильнее и очень сильно – срезание. Повреждение тканей стимулирует поглощение кислорода по трем причинам: 1) из-за быстрого окисления соединений, выходящих из вакуолей поврежденных клеток и становящихся доступными для окисляющих ферментов; 2) в связи с увеличением количества субстратов для дыхания; 3) вследствие активации процессов восстановления. Водоемы всегда беднее кислородом, чем атмосфера. В самых благоприятных условиях (при 0°) в 1 л воды может раствориться не более 14,7 мг О2 (1% объемной концентрации). С повышением температуры этот показатель падает, и при +25°, например, растворимость кислорода уже в два раза меньше. Кроме того, в погруженные в воду органы кислород проникает только путем диффузии через оболочки клеток (там нет устьиц), а это происходит очень медленно (скорость диффузии кислорода в воде в 104 раз меньше, чем в воздухе). Ситуация усугубляется тем, что в воде кислород диффундирует медленнее углекислого газа. Поэтому для находящихся в почве и воде корней, семян и многих аэробных микроорганизмов кислород часто становится лимитирующим фактором, и его недостаток сказывается на жизни растений. Кислород также активно расходуется на дыхание микрофлоры при разложении в воде органики, причем при высоких температурах этот процесс ускоряется. Такое «биохимическое потребление кислорода» особенно характерно для стоячих вод, где накапливаются органические загрязнители или листовой опад. Иная ситуация с кислородом складывается в почвах, особенно заболоченных и уплотненных. Уже в верхних дециметрах содержание кислорода быстро падает, а при застое воды может снижаться до нескольких процентов. Поэтому в зоне корней недостаток его встречается довольно часто. Аэрация почвы. Гипоксия и аноксия В жизни растений большое значение имеет аэрация почвы. От нее зависит: – скорость проникновения в почву кислорода и удаления из нее углекислого газа, – интенсивность накопления анаэробно образующихся соединений (сероводорода, метана и др.), – образование микроорганизмами и потеря необходимых растениям веществ (например, нитратов). Газообмен необходим для поддержания в почве достаточной концентрации кислорода, поскольку без притока из атмосферы его запасы летом, например, могут быть исчерпаны за 20-100 ч. Аэрация обеспечивается системой почвенных пор, заливание которых препятствует проникновению кислорода в почву. Содержание кислорода в почвенном воздухе зависит от многих причин: – погодных условий (температуры, влажнсти). Влияние температуры и влажности но эта связь неоднозначно. Так, при оптимальной влажности с повышением температуры содержание 02 в почвенном воздухе падает, но в летнюю жару и засуху концентрации его в почве могут быть максимальны, – типа почвы и ее физико-химических свойств (объем порового пространства, размер пор, степень дренированности и др.), – биологических параметров (интенсивность дыхания почвенных организмов и корней), – типа угодий (пашня, лес, луг). В почвах нормального увлажнения содержание кислорода, как правило, уменьшается от верхних горизонтов к нижним. На пашне состав воздуха связан с агротехникой и возделываемой культурой. На паровом поле в почвенном воздухе кислорода становится больше, а углекислого газа – меньше. Влияние нехватки кислорода в почве и избыточного увлажнения При нехватке кислорода: – изменяются отношения растений с почвенными организмами: увеличивается поражаемость паразитическими грибами, снижается активность микоризообразователей; – в заболоченной почве существенно изменяет деятельность почвенной микрофлоры. Так, в затопленном субстрате значительно повышается активность анаэробных аммонифицирующих (род Clostridium) и денитрифицирующих (Pseudomonas fluoresceins, Micrococcus denitrificans и др.) бактерий, что ведет к значительным потерям азота в почвах с плохой аэрацией; – усиливается токсическое действие некоторых продуктов разложения; – увеличивает кислотность почвы; – ускоряет оподзоливание и оглеение, образование вредных для растений закисных соединений; – замедленное разложение остатков организмов при дефиците кислорода также препятствует образованию гумуса и формированию благоприятной для корней комковатой структуры почвы. Избыточное увлажнение приводит к: – снижению содержания воздуха (кислорода) в почве; – заметному снижению интенсивность разложения гумусовых веществ аэробными микроорганизмами, что замедляет переход их в доступные для растений минеральные формы. Особенно ослабляется деятельность нитрифицирующих бактерий (Nitrosomonas, Nitrobacter – аэробы, активные лишь в хорошо аэрируемых незакисленных почвах, богатых органикой), обогащающих почву нитратами, используемыми растениями в азотном питании. – обеднению соединениями Са, К, Mg и основаниями, так как в составе их гумуса преобладают фульвокислоты, образующие с ними растворимые соли, мигрирующие вниз по почвенному профилю с просачивающейся влагой. Растения на таких почвах страдают от минерального голодания еще и потому, что даже если корни в них не отмирают, при нехватке кислорода они могут прекращать всасывание элементов минерального питания. Потребности корней в кислороде удовлетворяются не только за счет почвенного воздуха. Часть его диффундирует вниз от побегов. По межклетникам воздух, обогащенный выделяющимся при фотосинтезе кислородом, поступает из листьев в корни. Роль листьев в снабжении растений кислородом очень велика, поэтому при лишении света и дефолиации растения могут резко снижать урожайность и обнаруживать все симптомы анаэробного обмена. Показано, что воздух может выделяться из корневых окончаний в почву и аэрировать прикорневой участок, поддерживая на поверхности корней комплекс аэробных организмов. У разных растений этот процесс происходит с разной интенсивностью: количество кислорода, выделяемого единицей поверхности корней в единицу времени, у пушицы многоколосковой (Eriophorum polystachyori) в 13, у вахты (Menyanthes trifoliata) – в 16 раз больше, чем у молинии (Molinia). К.А. Куркин (1976) отмечает выдающуюся способность к аэрированию болотной почвы у тростника (Phragmites australis), обладающего сильно развитой воздухоносной тканью. Явления гипоксии (недостатка кислорода) и аноксии (отсутствия кислорода) широко распространены в природе. Очень часто в условиях кислородной недостаточности оказываются озимые (рожь (Secale cereale), пшеница (Triticum aestivum)), ячмень (Hordeum), соя (Glycine), рис (Oryza sativa), хлопчатник (Gossypium), некоторые древесные растения (ель (Picea), сосна (Pinus), береза (Betula), фруктовые деревья) и др. Одни растения при переувлажнении погибают очень быстро, другие способны переносить даже затопление и связанную с ним аноксию гораздо дольше. Однако все растения являются аэробами, поэтому в условиях аноксии они могут пребывать лишь временно. Воздействия гипоксии и аноксии на растения не идентичны. При гипоксии отключаются только системы, обладающие низким сродством к кислороду (например, окислительные системы микросом), а электрон-транспортная цепь митохондрий продолжает работать, хотя и с ограничениями. При аноксии же электрон-транспортная цепь полностью отключается, если нет соединений, заменяющих кислород как акцептор электронов. Физиологические изменения растений при ослабленной аэрации почвы: – усиление расхода растениями углеводов; – некоторых видов корни переходят к анаэробному дыханию; – повышается проницаемость клеточных мембран; – снижается рН клеточного сока; – снижается транспирация и скорость получения воды из почвы; – тормозится поглощение и передвижение воды и минеральных веществ из корней в листья; – в условиях резкого нарушения воздушного режима почвы может прекратиться снабжение корней продуктами фотосинтеза; – корни, которые обычно выделяют лишь углекислый газ, начинают отдавать ряд веществ (в иных условиях их выделение не отмечается). Так, из корней обратно в почву поступают некоторые из ранее поглощенных веществ. В отдельных случаях в составе корневых выделений обнаруживаются токсичные для растений газы (Н2, СН4, SО2 и др.) и кислоты (уксусная, щавелевая и др.), которые связываются в почве с элементами минерального питания, делая их недоступными для растений; – анаэробиозу обычно сопутствует избыточное выделение корнями СО2 и накопление этанола; – в целом в результате комплекса внутриклеточных процессов в условиях анаэробиоза происходит общее угнетение роста и развития растений, снижение их продуктивности. Устойчивость растений к гипо- и аноксии Устойчивость растений к гипоксии связана прежде всего со способностью сохранять близкое к нормальному содержание кислорода в тканях. Этому способствуют разнообразные анато-морфологические и физиологические. В случае же недостаточности их (особенно для вентиляции корневых окончаний) при длительном затоплении или снижении содержания кислорода в воде растения используют специальные метаболические приспособления к функционированию при недостатке или отсутствии кислорода. В связи с этим школой А.Л. Курсанова в 1980-е гг. разработана концепция двух стратегий адаптации растений в условиях анаэробного стресса, ставшая общепризнанной в физиологии. Первая стратегия – истинная толерантность, или адаптация на молекулярном уровне, реализуемая в условиях полного исключения кислорода из среды обитания. Вторая – кажущаяся толерантность, или адаптация на уровне целого организма, при которой клетки не имеют механизма резистентности к аноксии и избегают анаэробиоза благодаря транспорту кислорода из аэрированных частей в органы, локализованные в бескислородной среде. С 1970-х гг. в экологической физиологии растений большое значение придается также стрессовым белкам, появляющимся у растений в условиях аноксии и исчезающим в аэробных условиях. Индукция их синтеза при анаэробном стрессе играет существенную роль в обеспечении защиты мембран, а блокировка их образования в отсутствие кислорода приводит к дестабилизации мембран митохондрий и других органелл, что ведет к гибели клеток. Большое функциональное значение специальных стрессовых белков в адаптации растений к аноксии было подтверждено в экспериментах с кукурузой (Zea mays). 2.3 Адаптация растений к недостатку кислорода Анатомо-морфологическая адаптация к недостатку кислорода В заболоченных почвах некоторые из видов растений приобрели способность образовывать кочки и размещать при этом корни вне анаэробных условий. Так на сырых лугах ведут себя многие осоки (Carex cespitosa, С. juncella и др.) и злаки, например луговик дернистый (Deschampsia cespitosa), вейник Лангсдорфа (Calamagrostis langsdorffii). Ряд болотных и водных видов развиваетаэренхиму – воздухоносную ткань с увеличенными межклетниками и полостями, протягивающимися из надземных или надводных частей растений в органы, находящиеся в заболоченной почве или воде. Иногда происходит разрастание рыхлых тканей в основании стебля. Если такая ткань находится снаружи органа, то он приобретает белый цвет. По Г. Элленбергу (1978), хорошо развитую аэренхиму в корнях имеют более 13% сосудистых растений Центральной Европы. Воздухоносная ткань характерна и для видов, приспособленных к переменному увлажнению. У них также формируется толстостенная опробковевшая экзодерма, что является приспособлением к периодическому пребыванию корней в слабо- увлажненной почве. К этой группе растений относятся молиния (Molinia), хвощ полевой (Equisetum arvense), осока высокая (Carex elata), кровохлебка лекарственная (Sanguisorba officinalis), сухопутная форма горца земноводного (Polygonum amphibium). Некоторые растения хорошо аэрируемых почв формируют аэренхиму, оказав шись при недостаточной аэрации. Это свойственно даже ксерофильному кострецу прямому (Bromopsis erecta), а также мезофитам – райграсу высокому (Arrhenatherum elatius), еже сборной (Dactylis glomerata), овсянице луговой (Festuca pratensis) и др. При ухудшении аэрации многие виды образуют придаточные корни с более развитой аэренхимой. Для растений мангровых зарослей характерныпневматофоры (дыхательные корни). Во время прилива над водой возвышаются только кроны мангровых деревьев, а во время отлива обнажаются стволы и система особых корней – ходульных и дыхательных. Пневматофоры имеют возвышающиеся над субстратом части с отверстиями, через которые воздух проступает в воздушные полости и проходит в подземные органы растений. Дыхательные корни могут быть разного типа. У одних видов (Bruguiera) они коленчато выступают из ила. У других корни змеевидные, ползучие, и дыхательными отверстиями снабжен их верхний утолщенный край (например, у Xylocarpus). У третьих в результате отрицательного геотропизма (роста вверх, из земли) корни вертикально выходят из полужидкого субстрата (Avicennia, Sonneratia), а у Rhizophora их функцию берут на себя выступающие из воды части ходульных корней. Кроме мангровых растений дыхательные корни развивают некоторые пальмы, панданусы (Pandanus), сахарный тростник (Saccharum officinarum) и другие растения в почвах с недостаточной аэрацией. С недостатком кислорода в сырых почвах связывают иногда и образованиедосковидных корней у ряда тропических деревьев. Полагают, что плохая аэрация почв дождевых тропических лесов препятствует вторичному приросту древесины на внутренних сторонах боковых корней. Досковидные корни, развивающиеся у некоторых видов в нижних частях ствола, также служат для укрепления в почве крупных растений. Эти образования в виде крыльев отходят от ствола начинаясь у некоторых видов на высоте около 8 м. Образование досковидных корней обусловлено генетически. Они часто встречаются у некоторых видов фикусов, растений из семейств Sterculiaceae, Bignoniaceae и др. Из анатомо-морфологических изменений, вызываемых недостатком аэрации почвы, отмечается: – утончение клеточных стенок в корнях, – ослабление корневого ветвления и образования корневых волосков, – формирование придаточных корней в основании стебля. При этом ризосфера обычно имеет меньший объем, корни укорачиваются, занимают более поверхностное положение, масса надземной части растения снижается, площадь листьев и содержание в них хлорофилла падают, иногда наступает хлороз. У водных растений для максимального поступления кислорода из воды обычно увеличивается поверхность подводных органов. Это часто достигается рассечением листьев на многочисленные доли, их сильным удлинением и истончением. Метаболические приспособления растений к недостатку кислорода связаны с различными звеньями обмена веществ, среди них ведущие – адаптации дыхания. У устойчивых к гипоксии растений наблюдается: – низкая интенсивность дыхания даже при нормальной аэрации; – другое соотношение разных путей дыхания. Эти перестройки нужны для образования достаточного количества высокоэнергетических соединений и веществ для осуществления биосинтеза. При гипоксии критично не столько образование восстановителей, сколько координация их генерирования и реокисления. Основным путем окисления глюкозы в анаэробных условиях является гликолиз. Однако скорость переключения на него и динамика у разных по устойчивости к гипоксии растений различаются: – у неустойчивых увеличение доли и активности гликолиза резко возрастает уже в первые часы анаэробиоза, но оказывается кратковременным, – у приспособленных растений этот процесс менее интенсивен и более продолжителен, что обеспечивает более надежную поставку энергии. – для поддержания высокой скорости гликолиза необходимо непрерывное реокисление восстанавливающихся коферментов. При аноксии, когда электрон-транспортная цепь митохондрий отключена, окисление НАДН может происходить в конечных реакциях молочнокислого и спиртового брожения, но при этом накапливаются токсичные продукты этих процессов, особенно этиловый спирт: ü приспособленные к гипоксии растения накапливают его меньше. Этанол выделяется в ризосферу, что понижает его содержание в тканях устойчивых растений. Меньшее накопление этанола в тканях устойчивых растений связано и с его частичной переработкой; ü У неустойчивых видов, несмотря на более интенсивное выделение спирта, его концентрация нарастает в тканях и ризосфере, что дополнительно угнетает организм. У неустойчивых растений кратковременное возрастание доли пентозофосфатного пути происходит после увеличения доли гликолиза, а у устойчивых – сразу после ограничения доступа кислорода. Приспособительное значение этого процесса: – для неустойчивых растений незначительно, как и быстрое повышение активности гликолиза, так как это тормозит метаболизм и ставит их на грань гибели; – у приспособленных растений играет положительную роль, так как он определяет главным образом поставку восстановителя и соединений для биосинтеза. Несмотря на то что при аноксии электрон-транспортная сеть мито хондрий не работает, у приспособленных растений ее компоненты длительно сохраняют потенциальную активность. Поэтому использование при аноксии других акцепторов электронов у устойчивых растений тоже сохраняется дольше. Так, в анаэробных условиях при отсутствии кислорода как конечного акцептора электронов для растений оказываются важны процессы аноксического эндогенного окисления, в ходе которого электроны переносятся на такие вещества, как нитраты, – нитратное дыхание, двойные связи ненасыщенных соединений (жирные кислоты, каротиноиды). Перенос электронов на подобные акцепторы играет роль запасного выхода и способствует продлению жизни растений в бескислородных условиях. К метаболическим компенсаторным изменениям можно отнести также осуществляющийся в бескислородной среде синтез белков (большая часть их – ферменты анаэробного обмена), идущий несмотря на преобладающие процессы белкового распада.
3 Экологическое значение углекислого газа воздуха, поступление СО2 в растение и его регуляция Экологическое значение углекислого газа воздуха: – растения строят органические вещества своего тела за счет его ассимиляции при фотосинтезе. Для образования 1 г глюкозы необходимо поглотить 1,47 г (или 0,75 л) СО2 (учитывая его концентрацию, такое количество СО2 извлекается из 2500 л воздуха). – Углекислый газ имеет большое значение и в химическом изменении минеральной части почв. Почвенный раствор, насыщенный углекислотой, растворяет многие соединения почвы, в частности кальцит (СаСО3), доломит (CaCО3 ∙ MgCО3), магнезит (MgCО3), сидерит (FeCО3). Содержание углекислого газа в воздухе и его колебания Относительное постоянство содержания углекислоты в атмосфере поддерживается благодаря равновесию всех составляющих биогеохимического круговорота углерода и буферной роли Мирового океана. Однако концентрация С02 в воздухе постоянно увеличивается, возможно, благодаря интенсивному поступлению в атмосферу углерода, выведенного из круговорота в прежние геологические эпохи, прежде всего, в результате интенсивного сжигания ископаемого топлива. Намного больше углерода содержится в виде карбонатов в осадочных породах, живых и отмерших организмах, и даже в Мировом океане его в десятки раз больше, чем в атмосфере, что связано с высокой растворимостью СО2 в воде. Расчеты показывают, что к 2035 г. содержание СО2 в атмосфере может удвоиться и составить около 0,06%. Следствием этого может быть глобальное потепление климата, обусловленное так называемым «тепличным эффектом». Его связывают с тем, что углекислый газ «прозрачен» для основной части солнечного света, но задерживает тепловое (инфракрасное) излучение от нагретой Солнцем поверхности Земли. По некоторым расчетам, увеличение концентрации СО2 в атмосфере в 2 раза может привести к повышению температуры поверхности Земли на 2-3°, причем оно будет минимальным в тропической зоне и максимальным (8-11°) в высоких широтах. Сезонные колебания концентрации атмосферной углекислоты связаны с фотосинтезом и изменениями увлажнения, аэрации и температуры почвы, влияющими на почвенную микрофлору. Так, в высоких широтах после продолжительных дождей активность аэробных почвенных микроорганизмов падает, а содержание углекислоты в приземном слое воздуха снижается. В аридных областях угнетение почвенной микрофлоры и уменьшение содержания углекислоты в приземном слое воздуха вызывается засухой. Годичные колебания СО2 в воздухе особенно значительны в Северном полушарии, где преобладает суша. Весной в северных областях потребление углекислого газа развивающимися растениями значительно превышает его поступление из почвы. Суточные колебания содержания углекислоты более выражены, чем сезонные. В фитоценозах обычно наблюдается связанное с фотосинтезом дневное уменьшение концентрации СО2. Например, в лесу начало фотосинтеза утром обычно приводит к резкому падению его содержания. К полудню температура повышается, дыхание усиливается и количество углекислоты в кронах отчасти выравнивается. После захода солнца фотосинтез прекращается, начинает преобладать ды хание и концентрация СО2 вблизи почвы повышается. Изменение содержания углекислого газа происходит также при подъеме в горы. Например, на Памире на высоте 3800 м концентрация СО2 всего 0,012—0,020%. Источники атмосферной углекислоты Углекислота выделяется при извержениях вулканов (65 млрд т в год), сжигании горючих веществ, и некоторую прибавку дает разложение карбонатных горных пород. Однако главным источником углекислого газа в сообществах суши является дыхание почвы. Под ним понимают выделение СО2 эдафоном (совокупностью живых организмов почвы) в прилегающий к почве слой воздуха. В этом процессе на долю дыхания корней приходится до 10-30%, дыхание крупных животных очень мало, и основное количество СО2 образуется за счет микробиологической переработки остатков отмерших организмов (поэтому дыхание почвы является хорошим показателем интенсивности минерализации органического вещества почвы). Поступление углекислоты в растение Газообмен растения происходит путем диффузии, и на пути от атмосферного воздуха до хлоропластов двуокись углерода преодолевает ряд препятствий, определяющих в сумме диффузное сопротивление. В целом сопротивление транспорту углекислоты включает сопротивление пограничного слоя (воздушной пленки около листа), физиологически регулируемое устьичное сопротивление, диффузное сопротивление в системе межклетников, сопротивление при растворении и транспортировке СО2 в жидкой фазе клеточной стенки и в протоплазме, а также сопротивление в процессе карбоксилирования. Сопротивление пограничного слоя ниже, чем последующие устьичное и мезофильное сопротивления. У наземных растений СО2 проникает в лист через устьица (прохождение через клетки эпидермиса возможно только при очень высоких его концентрациях, в естественных условиях не встречающихся). Устьичное сопротивление – главное препятствие для свободного поступления углекислоты в растение. Закрытые устьица практически полностью перекрывают поток СО2 в лист. При недостатке света, когда устьичные щели суживаются, устьичное сопротивление становится фактором, ограничивающим диффузию СО2. При широко раскрытых устьицах это сопротивление ниже сопротивления мезофилла. Прошедшая через устьичную щель углекислота попадает в межклетники, где пополняется СО2, выделившимся при дыхании клеток. Сопротивление диффузии в системе межклетников зависит от строения листа. Обычно газопроводность теневых листьев выше, чем световых, а в мягких листьях она лучше, чем в жестких фотосинтезирующих органах. В клеточных стенках углекислый газ растворяется, и этот процесс тормозит диффузию. Внутри клетки растворенная углекислота очень медленно (по сравнению с газообразной фазой) движется к хлоропластам, где присоединяется к акцептору. Карбоксилирование может отставать от световых реакций фотосинтеза и лимитировать скорость этого процесса в целом. Сопротивление карбоксилирования зависит от эффективности работы ферментов – карбоксилаз, отличающихся в разных группах растений. Так как ФЕП-карбоксилаза катализирует связывание углекислоты намного эффективнее, чем РДФ-карбоксилаза, сопротивление карбоксилирования у С4-видов значительно меньше, чем у С3-растений. Устьичная регуляция поступления СО2 Изменяя ширину устьичных щелей, растение контролирует поступление углекислоты и одновременно идущую потерю воды. Количество, распределение, размеры, форма, подвижность устьиц видоспецифичны, но в некоторой степени изменчивы в зависимости от условий местообитаний и индивидуальных особенностей растений. Для поступления углекислого газа в растение решающее значение имеет площадь отверстий устьиц. С ее уменьшением устьичное сопротивление диффузии возрастает по экспоненте. У большинства растений от верстия устьиц занимают 0,5-1,5% поверхности листа. Максимальная диффузия газов в лист достигается при максимальном открытии устьиц, которое зависит от формы замыкающих клеток и осо бенностей строения их оболочек. Степень открытости устьиц зависит от состояния растения и внешней среды. Из экологических факторов наибо лее значимы свет, температура, влажность воздуха и водообеспеченность. Из внутренних факторов сильнее всего влияют на поступление углекис лоты парциальное давление в межклетниках, гидратация растения, баланс ионов и фитогормонов. Из гормонов гибберелиновая кислота и цитоки нины способствуют открыванию устьиц, а абсцизовая кислота – закрыванию. На состояние устьиц также влияют возраст листьев, стадия развития растения и эндогенные суточные ритмы. При хорошем водоснабжении устьица открываются тем шире, чем интенсивнее освещение – фотоактивное открывание. Оно определяется не только прямым, но и косвенным действием света (через расход СО2 на свету при фотосинтезе). Хемоактивная регуляции устьичных движений связана с содержанием углекислоты в воздухе. Так, при снижении ее содержания ниже критического значения они открываются и в темноте. Реакцию устьиц индуцируют и другие вещества (SО2, озон, сульфат алюминия и др.). На состояние устьиц влияют также некоторые токсины паразитов (например, выделения фитопатогенного гриба Helminthosporium maydis), а также вещества, применяемые для борьбы с ними (например, фунгицид фенилмеркурацетат). Температура влияет на скорость открывания устьиц через выработку энергии для их движения. При более высокой (но неэкстремальной) температуре энергии у клетки больше и устьица открываются быстрее. При сверхоптимальных температурах устьица также открываются не полностью, однако в жару (+40-45°) у некоторых видов ширина устьиц вновь достигает максимума. Важный фактор регуляции состояния устьиц – водообеспеченность. Устьичная щель закрывается при потере воды и падении тургора замыкающих клеток. В сухом воздухе реакция закрывания устьиц наступит раньше, чем растение начнет испытывать серьезный недостаток воды. Ветер, сдувающий водяной пар с листа, усиливает этот эффект.
Физиологический механизм движений замыкающих клеток связан с изменением тургора. В расслабленном состоянии устьичные щели закрыты, и их открывание требует затрат энергии. Тургор может быть повышен активным транспортированием ионов (К+, РО43-), изменением проницаемости и ферментативными процессами. При этом существенны запас АТФ, процессы карбоксилирования, ферментативные превращения крахмала и сахаров. 4 Непостоянные компоненты воздуха. Проникновение токсичных газов в клетки растений и их действие Непостоянные компоненты воздуха Непостоянные компоненты воздуха могут содержаться в атмосфере разных местообитаний в разных количествах. Примеси в воздухе имеют большое значение для растений, особенно в промышленных центрах. Наиболее вредными компонентами промышленных газов являются оксиды серы, азота, соединения фтора и хлора. Основной источник дымовых газов – сжигаемое топливо. Среди атмосферных загрязнителей для растений особенно опасны продукты сгорания, содержащие сернистый ангидрид (SО2), который вызывает разрушение хлорофилла, недоразвитие пыльцы, нарушает работу проводящей системы и др. Выхлопные газы содержат свинец, токсично действующий на растения вблизи автодорог. Соединения металлов (меди, цинка, кобальта, никеля, кадмия и др.) неблагоприятно влияют на растения вблизи металлургических предприятий, поступая в них из почвы и воздуха. Газообразные примеси абсорбируются почвой и подстилкой, соединяются с водой и могут образовывать кислоты. Они повышают кислотность почвы и уменьшают насыщенность ее основаниями, что способствует снижению биохимической и микробиологической активности почвы. Кислоты могут переводить в растворимую форму вещества, токсичные для растений. Пыль, содержащаяся в воздухе, влияет на растения. Она может снижать прозрачность воздуха и, уменьшая освещенность, ограничивать углеродное питание растений. Осевшие на поверхности листьев пылевые частицы действуют как экран, понижающий доступ ФАР и усиливающий поглощение тепла. Они также могут привести к закупориванию устьичных щелей. Все это ухудшает условия фотосинтеза и водообмена. Особенно вредна цементная пыль, во влажной среде образующая на листьях плотную корку. Для растений токсичны и взвешенные в воздухе частицы моющих средств. Пыль и дымовые газы могут существенно модифицировать микроклимат местообитаний. В промышленных зонах нередко повышается температура и падает влажность воздуха. В аридных районах нередко случаются пыльные бури, способствующие сильному иссушению воздуха. Проникновение токсичных газов в клетки растений Многие токсичные вещества при газообмене проникают в растение из воздуха, с дождем, при осаждении тумана и пыли. Основная часть приносимых по воздуху загрязняющих веществ перехватывается листьями. Они адсорбируются и выводятся из обмена кутикулой, клеточными стенками, срединными пластинками, вакуолями, которые могут стать их хранилищами. Первая преграда на пути проникновения токсинов из ат мосферы в растение — кутикула. Так, на поверхности листьев концентрируется существенная часть соединений свинца из выхлопных газов. Кутикула представляет собой гетерогенную мембрану, состоящую из тонкого внешнего слоя, препятствующего попаданию веществ из внешней среды, и внутреннего сорбционного слоя. Благодаря им кутикула эффективно выполняет двойную роль барьера и поглотительной емкости для химических загрязнителей. Вещества кутикулы способны ковалентно связывать химические соединения с карбоксильными заместителями, образуя сложные эфиры. Ковалентно связанные в кутикуле химические соединения сохраняются в растении, не подвергаясь детоксикации, но могут высвободиться в пищеварительном тракте животных и человека или при разложении почвенной подстилки. Кутикулярный барьер листьев обычно не пропускает токсичные газы в растение. Поэтому основная масса их попадает в листья при газообмене через открытые устьица и чечевички. При этом происходит нарушение регуляторной функции замыкающих клеток, токсичные вещества накапливаются в межклетниках и через клеточные оболочки проникают к клеткам мезофилла. Многие токсичные газы растворяются водой, содержащейся в клеточной оболочке (она составляет около 60% массы листа), реагируют с ней и образуют кислоты. При этом содержание подвижной воды в клетке падает, что снижает скорость движения цитоплазмы, нарушает внутриклеточный транспорт и приводит к плазмолизу. Чем больше растворимость загрязнителя в воде, тем выше скорость его поглощения и сильнее воздействие на организм. Газы, об
|
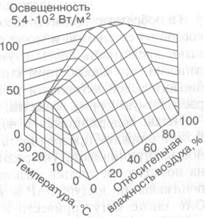 Совокупное влияние внешних факторов обычно обусловливает не максимальную, а среднюю степень открытия устьиц. Полное закрытие устьичных щелей вызывают тоже только экстремально неблагоприятные условия.
Совокупное влияние внешних факторов обычно обусловливает не максимальную, а среднюю степень открытия устьиц. Полное закрытие устьичных щелей вызывают тоже только экстремально неблагоприятные условия.


