Высвобождение медиатора
Микроморфология концевой пластинки. Существующие представления о структуре концевой пластинки отражены на рис. 3.13. Характерная особенность пресинаптического окончания мотонейронаскопления в нем (кластеры) «синаптических» пузырьков. Напротив них постсинаптическая мембрана образует глубокие складки. Каждой из них соответствует активная зона пресинаптической мембраны-желобок на ее внутренней поверхности, вдоль обеих сторон которого располагаются в ряд синаптические пузырьки. Некоторые из них открыты наружу, в синаптическую щель. Очевидно, активные зоны и ассоциированные с ними пузырьки следует рассматривать как аппарат, специализированный для экзоцитоза (с. 17), т.е. для выброса содержимого этих пузырьков в синаптическую щель. Биохимическими методами показано, что они содержат высокие концентрации ацетилхолина, а также белки и нуклеотиды. Таким образом, в активных зонах
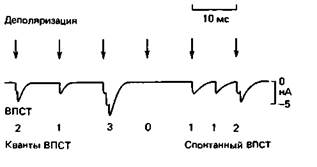
ацетилхолин высвобождается из окончания мотонейрона в виде накопленных пузырьками порций. «Квантовая» природа потенциала концевой пластинки. Поскольку ацетилхолин высвобождается приблизительно одинаковыми порциями, соответствующими объему пузырька, постсинаптический ток (ВПСТ) должен состоять из мелких «субъединиц». Их можно наблюдать, регистрируя с помощью метода локальной фиксации потенциала «пэтч-кламп» (см. рис. 2.11); синаптические токи в микроучастке концевой пластинки длиной несколько микрометров. Во время стимуляции мотонейрона амплитуда ВПСТ явно представляет собой кратное количество этих субъединиц; на рис. 3.14 показаны последовательно 2, 1, 3, 0... его «квантов». В отсутствие стимуляции нейрона они могут появляться спонтанно. Количество квантов на один стимул варьирует стохастически, биномиально рас- 62 ЧАСТЬ I. ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ пределяясь около некоторого среднего значения. Практически нет сомнений, что каждый такой «квант» тока соответствует порции ацетилхолина из одного пузырька, достигающей путем диффузии постсинаптических рецепторов и вызывающей открывание ионных каналов. Пузырек содержит несколько десятков тысяч молекул ацетилхолина. Длина пресинаптического окончания концевой пластинки составляет более I мм; из него в ответ на один потенциал действия двигательного аксона высвобождается несколько сот «квантов», неразличимых индивидуально в суммарной реакции. Однако между потенциалами концевой пластинки, достигающими примерно 40 мВ (рис. 3.2), можно зарегистрировать спонтанные сдвиги потенциала амплитудой менее 1 мВ, обусловленные спонтанным высвобождением «квантов» медиатора из открывающихся наружу синаптических пузырьков. Медиаторы запасаются в пузырьках и высвобождаются из них не только в нервно-мышечном, но и во всех других известных химических синапсах. Например, «квантовые» токи на рис. 3.12 вызваны таким высвобождением глутамата. Пузырьки могут содержать различные медиаторы, перечисленные на рис. 3.7, но обычно только по одному в каждом синапсе. Впрочем, иногда наряду с классическим медиатором (например, ГАМК) в пузырьках находится и пептид с модулирующим действием (с. 54). Высвобождение квантов медиатора. Потенциал действия пресинаптического окончания ведет к почти синхронному (с небольшой синаптической задержкой) высвобождению квантов медиатора, которое приводит к генерированию потенциала (например. ВПСП) в постсинаптической мембране. Временные соотношения этих процессов показаны на рис. 3.15 для гигантского синапса кальмара, где можно зарегистрировать как пре-, так и постсинаптические изменения потенциала и токи. Кроме потенциала действия, постсинаптических токов и потенциалов на рисунке представлено поступление Са2+ в пресинаптическое окончание, сопровождающее Na+- и К +-токи во время деполяризации (см. с. 38). Этот входящий Ca2 + -ток играет ключевую роль в квантовом высвобождении. Уже давно было известно, что химическая синаптическая передача нарушается при значительном снижении внеклеточной концентрации Cα2 +, [Са2 + ]0. Этот эффект примерно пропорпионален четвертой степени [Са2 + ]0 [12], следовательно, для высвобождения одного кванта медиатора требуется реакция четырех ионов Ca с активатором на внутренней стороне пресинаптической мембраны. Однако действие активатора зависит, по-видимому, еше и от потенциала, т.е. даже при достаточно высокой внутриклеточной концентрации Са2+,[Са2+]i синхронное высвобождение медиатора требует деполяризации мембраны [31]. Можно предполагать, что она влияет на акти-
ГЛАВА 3. МЕЖКЛЕТОЧНАЯ ПЕРЕДАЧА ВОЗБУЖДЕНИЯ 63 ватор примерно таким же образом, как и на молекулу ионного канала (рис. 2.12-2.15). Следовательно, пресинаптические активные зоны с их участками связывания пузырьков и мембранными белками («частицами») (рис. 3.13) должны представлять собой аппарат для быстрого регулирования экзоцитоза посредством деполяризации мембраны и повышения [Са2+];. Рост [Ca2+]j, возможно, влияет на сократительные элементы цитоскелета (см. с. 20, рис. 1.13) или инициирует фосфорилирование функциональных белков (рис. 1.16). Синаптическое облегчение. В свете представлений о квантовом высвобождении рассмотрим теперь синаптический механизм, сравнимый по своему значению с суммацией и торможением,— синаптическое облегчение. Этот процесс иллюстрирует рис. 3.16, А Когда окончание нейрона начинают раздражать с частотой 20 Гц, ВПСП, генерируемый в ответ на первый стимул, едва различим, но по мере продолжения стимуляции его амплитуда постепенно нарастает. Таким образом, ритмическая активация повышает эффективность синаптической передачи. Если частоту стимулов увеличить вдвое (рис. 3.16,^4, нижняя запись), эффект облегчения усиливается. Более того, благодаря значительному сближению ВПСП между собой происходит и их суммация (рис. 3.10) с повышением фонового уровня деполяризации, от которого начинается каждый ВПСП. Регистрация синаптических токов показывает, что при облегчении амплитуда ВПСТ' возрастает. Как иллюстрирует рис. 3.16, Б, во время торможения увеличивается среднее число квантов, высвобождаемых в ответ на каждый стимул. Наибольшее облегчение наблюдается, когда очередной стимул поступает через несколько миллисекунд после предыдущего; оно затухает с постоянной времени порядка 50 мс. Такое облегчение -пре синоптический процесс, поскольку при нем увеличивается вероятность высвобождения квантов медиатора. Большинство авторов полагают, что это обусловлено «остаточным кальцием». Во время деполяризации окончания в него входят ионы кальция и [Ca2 + ]j возрастает (рис. 3.15). Затем [Са2 + ]; возвращается к уровню покоя за счет процессов транспорта и обмена. Однако пока [Ca2+]j превышает этот уровень, при каждой следующей деполяризации повышение [Са2 + ]; начинается от более высокого значения, чем предыдущее, и в результате [Са2 + ]; постоянно нарастает. Так как высвобождение медиатора пропорционально, скажем, четвертой степени [Ca2 + ]i, даже относительно небольшой прирост этой концентрации обеспечивает существенное облегчение [20, 30]. Разным синапсам свойственна неодинаковая его степень. Ярко выраженное облегчение, как, например, на рис. 3.16, особенно характерно для цент-
ральных синапсов; здесь одиночный пресинаптический потенциал действия едва ли вызовет высвобождение даже одного кванта, тогда как несколько импульсов, быстро следующих друг за другом, гораздо более эффективны. Облегчение составляет своего рода «память» нервного окончания: в течение нескольких сотен миллисекунд в нем сохраняется след от предыдущего события. Известны и синапсы, в которых облегчение сохраняется минутами. Вполне вероятно, что синаптическое облегчение - первый этап формирования краткосрочной памяти, на основе которой может затем развиваться долгосрочная (см. гл. 6). 64 ЧАСТЬ I. ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ Облегчение, вызываемое относительно длинными сериями потенциалов действия, называют также синаптической потенциацией. Нарастание ВПСП, иллюстрируемое рис. 3.16, А, относится к тетанической потенциации. Сильное облегчение, сохраняющееся после стимуляции, а если она очень продолжительная, не исчезающее в течение нескольких часов, представляет собой посттетаническую потенциацию. Вероятно, во время таких длинных серий стимулов в пресинаптическом окончании одновременно с [Са2+]| повышаются концентрации и других ионов, в частности Na +. Еще один возможный эффект активации окончания, требующий участия внутриклеточного посредника,-подготовка пузырьков к высвобождению медиатора, т. е. их мобилизация [27]. Длительное высокочастотное возбуждение пресинаптических окончаний может в конечном итоге привести к состоянию, противоположному облегчению, - депрессии,когда количество квантов медиатора, высвобождаемых в ответ на потенциал действия, снижается. Истинные механизмы этого неясны. Одна из возможных причин - истощение запаса синаптических пузырьков с медиатором. Кроме того, следует иметь в виду, что нервные окончания, перед тем как образовать синапсы, обычно разветвляются, а точки ветвления - слабое звено в распространении потенциала действия. То же самое количество тока, которое при возбуждении входит в волокно перед точкой ветвления, должно за ней деполяризовать уже два волокна. Поэтому во время высокочастотного разряда проведение возбуждения в аксонные ветви часто блокируется. Это также проявляется как депрессия синаптической передачи. Возможно, депрессия, вызываемая ритмической активацией синаптического пути, в виде «привыкания» (термин, заимствованный из физиологии поведения) составляет основу процессов научения и памяти.
|