Головна сторінка Випадкова сторінка
КАТЕГОРІЇ:
АвтомобіліБіологіяБудівництвоВідпочинок і туризмГеографіяДім і садЕкологіяЕкономікаЕлектронікаІноземні мовиІнформатикаІншеІсторіяКультураЛітератураМатематикаМедицинаМеталлургіяМеханікаОсвітаОхорона праціПедагогікаПолітикаПравоПсихологіяРелігіяСоціологіяСпортФізикаФілософіяФінансиХімія
Photosynthesis and the Chloroplast
Дата добавления: 2014-11-12; просмотров: 593
|
|
Нет ничего труднее, чем завершение книги. С каждой страницей, приближающей к концу, умножаются сомнения: обо всем ли написал и так ли, как нужно. Воистину, по словам Гёте: «Alles war gesagt, doch alles bleibt zu sagen!» – «Все было сказано, лишь все сказать остается!» К счастью, на сей раз положение лучше. Вариант книги в 1980 году был опубликован популярным журналом «Наука и жизнь»; как водится, в редакцию пошли письма имеющих досуг читателей, и теперь ясно, что я недоучел в работе. Разумеется, далеко не все письма могли быть использованы. Редакция предпочла сама отвечать на те письма, которые не содержа ли ничего, кроме брани. Однако и среди тех, что были мне направлены для ознакомления, находились такие, которые до сих пор меня ставят в тупик.
Биология наряду с науками, являющимися ее практическим приложением (медициной, агробиологией и т. д.), в представлении многих моих корреспондентов кажется принципиально иной отраслью знания, чем, скажем, машиностроение или теория атомного ядра. Вряд ли кому‑нибудь придет в голову усомниться в фактах и выводах, изложенных, к примеру, в статье об управляемом термоядерном синтезе. А вот биология, особенно общая – здесь «каждый мнит себя стратегом, видя бой со стороны». И вот бедняга‑автор, рискнувший ступить на трудную стезю популяризации, получает письма об установлении связи с «космической цивилизацией Эридана Тулона Элсилон», о «всеобъемлющем законе энергии разности потенциалов» и т. д. и т. д. В эпитетах корреспонденты не стесняются, слова «идеализм», «недомыслие», «безграмотность», «узкая щель мирозрения» (так! – Б. М.) на общем фоне звучат чуть ли комплиментами.
Не скажу, чтобы подобные письма было приятно читать. Однако и из них можно почерпнуть полезное, чтобы еще раз остановиться на тех вопросах, о которых мои корреспонденты имеют ложное представление.
Любопытно, что читателей, насколько могу судить, в первую очередь интересуют именно философские проблемы, которые возникают при попытках аксиоматизировать биологию. Много говорится, например, о законе причинности. По‑видимому, все довольно легко согласились с тем, что учение Ламарка этот закон нарушает и мы должны сделать выбор: или мираж направленной приспособительной изменчивости, или же сохранение одного из основных принципов современного естествознания. Однако для некоторых мираж оказывается предпочтительнее закона причинности.
В некоторых письмах категорично утверждается: закон причинности устарел как для живой природы, так и для неживой. Один мой корреспондент упоминает тахионы, которые будто бы движутся со сверхсветовой скоростью, и настоятельно рекомендует мне прочесть популярные брошюры на эту тему. (Невольно вспоминается фарадеевское: «Популярные книги ничему научить не могут». Начинать можно с них, но завершать обучение, если дело касается физики, например, лучше все‑таки вузовским курсом Ландау и Лифшица.) Тахионы – частицы гипотетические, экспериментально они не обнаружены, реальное существование их не доказано, и поэтому опровергать с их помощью принцип причинности, мягко выражаясь, преждевременно.[10]
Не меньшее раздражение вызвало у ряда читателей то, что я подчеркиваю везде функциональную подоплеку аксиом, не связывая жизнь с каким‑либо химическим веществом. В этом усматривали идеализм и «богословную мистику». Однако вынужден еще раз подчеркнуть: да, в конкретных условиях нашей Земли материальным субстратом генетических программ оказались нуклеиновые кислоты, а функциональным, эффекторным субстратом – белки. Но из этого отнюдь не следует, что везде во Вселенной дело обстоит точно так же. Даже в условиях, приближающихся к земным, в том интервале давлений и температур, в котором существует жидкая вода, возможны материальные субстраты жизни, которые нам сейчас показались бы невероятными. Тем более это относится к планетам, покрытым морями, например, из жидкого метана и аммиака, даже из жидкой серы. Как бы нам ни казалось невозможным появление и эволюция жизни в таких условиях, вероятность этого во Вселенной нельзя отрицать с порога. Сможем ли мы узнать жизнь, столь непохожую на земную, или же будем искать в космосе только собственное отражение? Не придется ли нам тогда искать слишком долго?
Когда космонавты будущего обнаружат где‑то во Вселенной структуру, пусть любого, самого фантастического вида и строения, если эта структура может поддерживать свою целостность и воспроизводить себе подобных согласно вложенной в нее программе, реплицирующейся матричным способом, они должны признать ее живой. Дело даже не в будущих космонавтах, – понять, что такое жизнь, мы должны уже сейчас. Ибо это означает познание самого себя.
Пожалуй, наибольшее разнообразие можно было обнаружить в определениях понятия жизнь». Каких только формулировок не предлагают в дискуссиях и в письмах! Как правило, они не выдерживают критики, мягко скажем, из‑за недостаточной информированности авторов. Приведу лишь один пример, из самых характерных. Один автор чрезвычайно агрессивно отстаивающий свою точку зрения, доказывает, что в понятие жизни должен входить активный поиск и захват источников энергии. По его мнению, жизнь начинается с амебы: ведь она активно захватывает пищевые частицы. А вот бактерии, грибы, да и растения – неживые.
Ясно, автор не подозревал о существовании хотя бы бактерии бделловибрион бактериоворус (что: означает пиявкообразный вибрион, пожирающий бактерий). Этот маленький вибрион нападает на других бактерий, причем движется с огромной скоростью – до ста своих длин в секунду! Если бы человек в своих масштабах мог двигаться так быстро, он пробегал бы стометровку за долю секунды. Бделловибрион ударяет бактерию‑жертву с такой силой, что она по инерции проходит немалое расстояние. А крошечный хищник тем временем стремительно просверливает клеточную стенку жертвы (скорость вращения свыше ста оборотов в секунду), за несколько секунд проникает в пространство между стенкой и мембраной, и через четыре часа из «съеденной» изнутри бактерии выходят новые, молодые вибрионы.
Если уж это не активный поиск источника энергии… Кстати, бделловибрионы – не единичный каприз природы. Хищные бактерии – мощный фактор очищения воды. Не слыхивал мой оппонент и о хищных грибах, обитающих в почве, мицелий которых, переползая с места на место, захватывает мелких животных; говорят уже об их применении для борьбы с почвенными нематодами – вредителями сельского хозяйства.
Но неужели мой оппонент ничего не знал о насекомоядных растениях, хотя бы о всем известной росянке, которую изучил еще Чарльз Дарвин? Или о часто встречающейся в наших водоемах пузырчатке: достаточно мелкому животному прикоснуться к чувствительному «волоску», запоры ловчего пузырька резко открываются, и ток воды затягивает жертву внутрь, где она и переваривается. Примерно так же захватывает добычу рыба – морской черт.
От растений, питающихся как животные, перейдем к животным, питающимся как растения. Симбиотические одноклеточные водоросли – зоохлореллы и зооксантеллы – синтезируют углеводы в клетках корненожек и гидр, губок и кораллов, плоских червей и моллюсков. О том, что такой способ питания (без активного поиска и захвата пищи) весьма эффективен, свидетельствуют хотя бы протянувшиеся на тысячи километров коралловые рифы. Ограничимся двумя примерами.
Осыхающая во время отлива поверхность песчаных пляжей Северного моря покрывается оливково‑зелеными пятнами. Это выползают на поверхность песка, под солнечные лучи маленькие плоские черви – конволюты. Тело их заполнено симбиотическими одноклеточными водорослями – зоохлореллами. За время отлива симбионты успевают насинтезировать углеводов и для себя, и для переваривающего их черня. Растение это или животное?
Гигантский двустворчатый моллюск коралловых рифов – тридакна, если никем не потревожена, открывает свои створки и выворачивает навстречу солнечным лучам оливково‑бурую мантию. Поверхность мантии как бы шагреневая. Она покрыта тысячами мелких выступов, заканчивающихся линзоподобными структурами из прозрачных, хрящеподобных (гиалиновых) клеток. Каждый такой бугорок – настоящий световод, собирающий солнечные лучи и передающий их внутрь ткани, к скоплениям симбиотических водорослей – зооксантелл. У тридакны недоразвита пищеварительная система, она живет главным образом за счет своих симбионтов, потребляемых клетками‑фагоцитами. Везде, где активное питание можно заменить пассивным, за счет утилизации энергии солнечных лучей, животные охотно делают это. И наоборот, там, где в почве мало азота, растения приобретают способность к хищничеству.
Мы видим, что построения нашего автора, строгого хулителя дарвиновской теории эволюции и современной генетики, рассыпаются как карточный домик при малейшем соприкосновении с фактами. Ибо это порождения – не постесняемся резкого слова – элементарного невежества. Вряд ли имело бы смысл останавливаться на явлениях подобного рода, если бы это невежество не было воинствующим. Слов нет, недостаток информации никогда не приносил пользы, однако воинствующее невежество может принести огромный вред, чему мы видим немало примеров в прошлом. Ведь тот же автор и его единомышленники голословно отрицают роль ДНК в явлениях наследственности. И это сейчас, когда успехи генной инженерии убедительно показывают, что до широкого внедрения в практику методов направленного изменения наследственности остаются считанные годы.
Генная инженерия может произвести подлинную революцию в медицине и сельском хозяйстве, промышленности и охране природы. (В то же время не будем закрывать глаза: в руках недобросовестных людей она может быть не менее страшной, чем ядерное оружие.) Зачеркивать достижения биологии последних лет – все равно что отрицать существование атомов в начале августа 1945 года[11]. И самое малое, что мы можем сделать, для того чтобы не отставать в развитии биологии, – решительно пресекать все попытки подобного, как писали Ильф и Петров, «головотяпства со взломом».
Но довольно об этом. Еще раз перечислю выдвинутые в этой книге в качестве аксиоматических положения, лежащие в основе жизни.
Именно:
1) живые организмы состоят из фенотипа и генотипа, то есть генетической программы;
2) генетические программы не возникают заново, а реплицируются матричным способом;
З) в процессе репликации неизбежны ошибки на микроуровне, случайные и непредсказуемые изменения генетических программ;
4) в процессе постройки фенотипа эти изменения многократно усиливаются, что делает возможным селекцию единичных квантовых событий на макроуровне.
Осмелюсь утверждать, что этих положений достаточно (как говорят математики: необходимо и достаточно) для объяснения всех феноменов живой природы – от возникновения первого сократимого волокна или первой молекулы фермента до становления мыслящего мозга. Я предвижу хор возмущенных голосов, основным лейтмотивом которого будет: «Нельзя сводить биологию к физике и химии!»
Но разве рассмотренные в этих очерках аксиомы характерны для неживой природы? Где же тут сведение? Появилось уже хлесткое определение «редукционизм» так называют направление, которое не прибегает для объяснения сущности жизни к аналогам «жизненной силы». Но, как я уже писал вначале, мы не сводим биологию к химии и физике. Мы выводим биологию из химии и физики. Это не редукционизм, а дедукционизм (различие в одной букве и во всем смысле термина).
Если уж говорить о редукционизме, то только в том смысле, что все эти положения прямо вытекают из положений физики и химии и только в этом понимании «сводятся» к ним. Ибо что значит – объяснить? Это и есть свести сложное явление к более простым, для нас уже понятным, объясненным. Утверждение, что это‑де отрицает «специфику жизненных явлений», по сути дела, повторяет анекдотическое определение жизни, цитированное мной раньше («живое состоит из живых объектов, неживое – из неживых»). Естествоиспытатель в наши дни обязан быть редукционистом, иначе он рискует остаться просто болтуном.
Вот, например, что пишет о редукционизме советский философ Р. С. Карпинская: «…если вопрос о редукционизме рассматривать в методологическом плане, то сущностью „сведения“ сложных биологических процессов к более простым является обнаружение на молекулярном (или ином, более простом по отношению к изучаемому) уровне таких фундаментальных характеристик, которые при их теоретическом обобщении позволят сформулировать некие абстрактные понятия, выступающие начальным пунктом движения познания „вверх“, ко все более сложным уровням биологической организации. Эти понятия должны работать на всех уровнях, наполняясь все более конкретным, все более богатым содержанием » (курсив мой. – Б. М.).
Прошу извинения у читателей за длинную цитату: на мой взгляд, в ней удачно подчеркивается необходимость неких абстрагированных заключений (которые я назвал аксиомами, но не в слове суть: назовите их постулатами, принципами, положениями, в конце концов, русский язык достаточно богат). Кроме того, формулируются условия, которым должны соответствовать наши аксиомы.
1. Хотя они (аксиомы) выводятся на молекулярном уровне (деление организма на генотип и фенотип, матрицирование генотипа, ошибки матрицирования, усиление ошибок, делающее их доступным отбору), с их помощью можно познать более сложные уровни биологической организации.
2. Они должны быть всеобщими, проявляться на всех уровнях и во всех случаях. Частные закономерности должны выводиться из них, как теоремы из аксиом. И включение в систему аксиом «лишней», не работающей на всех уровнях, такая же, если не худшая, ошибка, как включение неверной, не обоснованной фактами и противоречащей принципам физики и химии.
С этой точки зрения рассмотрим вопрос: достаточна ли наша система аксиом, не следует ли чего‑либо добавить к ней? Признаюсь, что для меня этот вопрос проклятый, я ломал голову над ним годами. Остановлюсь для примера на положении, которое чуть было не стало пятой аксиомой. Его можно было бы выразить двумя словами: генетическая рекомбинация.
Генетическая рекомбинация – обмен частями генетических программ – возникла, по‑видимому, на чрезвычайно ранней стадии становления жизни. У самых простейших организмов – бактериофагов – описан сложный процесс обмена кусками ДНК. После заражения бактериальной клетки фагом в ней образуется 30–50 копий молекул ДНК, реплицированных с материнской молекулы, впрыснутой в бактерию.
Дочерние молекулы могут спариваться, сближаться гомологичными частями; затем в зонах спаривания происходят разрывы нитей ДНК, обмен скрещенными участками и последующая сшивка. Все это очень напоминает хорошо известный генетикам перекрест хромосом у высших организмов – кроссинговер, при котором хромосомы обмениваются гомологичными частями. В результате, если в одной молекуле ДНК произошла одна мутация, а в другой – другая, они могут объединиться в генетической программе фага и совместно попасть под действие отбора.
Скорее всего, механизм этот развился из репарации, о которой мы здесь говорили. Репарационные ферменты – лигазы – сшивают разорванные жестким излучением или иным мутагеном нити ДНК, восстанавливают целостность генетической программы. А так как в одной клетке несколько десятков фаговых ДНК, то высока вероятность объединения частей разных молекул. Рекомбинация оказалась выгодной, допускающей объединение генетических изменений у разных организмов.
У бактерий перенос генетической информации осуществляется от клетки к клетке непосредственно кусками ДНК (трансформация), в результате изучения этого процесса и была доказана роль ДНК как субстрата наследственности. Часто гены переносятся от бактерии к бактерии фагами, фаг может прихватить кусок ДНК хозяина и передать его в новом поколении другому. Этот процесс называется трансдукцией. Обычно ее осуществляют плазмиды, «ручные» фаги, ставшие симбионтами бактерий и не убивающие хозяина интенсивным размножением. Из подобных механизмов у бактерий в конце концов развился оригинальный половой процесс конъюгация. При нем клетки соприкасаются, между ними образует цитоплазматический мостик по которому нить ДНК перетекает из одной бактерии в другую. Любопытно, что для передачи генного материала бактерия должна иметь внехромосомный генетический элемент, названный фактором F , который, возможно, потомок плазмиды. Не имеющие его штаммы (F – ) могут только принимать чужую ДНК (аналогия между самками и самцами у высших животных; кстати, «самцов» то есть имеющих фактор F , довольно мало: у кишечной палочки их в десять раз меньше, чем «самок»). Плазмида, ведущая происхождение от фага, может вообще стать частью бактериальной хромосомы, тогда она называется эписомой. Также и F – фактор, включаясь в геном бактерий, повышает их способность к конъюгации в тысячу раз.
Из школьного курса общей биологии вы должны знать, что генетическая рекомбинация у высших организмов гораздо сложнее. И у них есть кроссинговер. Но в отличие от бактерий геном у них настолько велик, что не может быть объединен в одной хромосоме.[12]
Ядерные организмы – эукариоты – имеют в клетках от двух (у малярийного плазмодия и лошадиной аскариды) до тысячи и более хромосом, «томов» генетических программ. Перед каждым делением клетки генетические программы реплицируются. Однако так бывает не всегда. Перед наступлением полового процесса происходит мейоз – редукционное деление. Хромосомы при нем не делятся, а расходятся по клеткам, из которых потом формируются половые. Так, у человека в норме 46 хромосом из них две половые, остальные 44 идентичны у обоих полов (аутосомы). 23 хромосомы человек получает от отца, 23 – от матери, а каково сочетание отцовских и материнских хромосом в яйцеклетке или спермии – это дело случая. Число сочетаний здесь равно двум в степени n – числу хромосом в гамете. Отсюда следует, что, например, у дрозофилы с ее четырьмя хромосомами в гамете число вариантов гамет 24 = 16, у кролика 222 = 4 385 000. Предоставляю читателям самим прикинуть число вариантов гамет у папоротника (количество хромосом в гамете 630). Слияние половых клеток восстанавливает прежнее количество хромосом возникает новая генетическая программах с которой организм входит в жизнь. При смене поколений эти программы рассыпаются чтобы заново возникли другие. Генетическая рекомбинация непрерывно тасует их, как карты, поставляя отбору поистине неисчерпаемый материал. Здесь есть и отрицательная сторона: какая‑нибудь сверхудачная комбинация хромосом бесследно исчезает в новом поколении, у гениальных родителей появляются заурядные дети.
По‑видимому, этот великий по простоте механизм рекомбинации возник в процессе эволюции только один раз. У человека и сосны, инфузории и дрожжевого грибка половые ядра, сливающиеся впоследствии, возникают одинаковым путем: ядро с нормальным набором хромосом не разделяясь реплицирует их, так что образуется четыре набора генетических программ. Затем клетка (или только ядро у инфузорий) дважды делится, получаются четыре клетки с половинным, гаплоидным набором. Часто, особенно при формировании яйцеклеток, три из них рассасываются, дальнейшее развитие суждено только одной. Лишь немногие панцирные жгутиковые имеют одноступенчатое редукционное деление; у них это, скорее всего, вторичное упрощение.
Так, может быть, объявим генетическую рекомбинацию непреложным законом живого, пятой аксиомой? Увы, дело обстоит не столь просто. Многие организмы потеряли способность к генетической рекомбинации и благоденствуют, мы не имеем права исключать их из мира живого. Таковы всем известные амебы и инфузории потерявшие способность образовывать половое ядро, все формы, размножающиеся партеногенетическим путем, без оплодотворения (а их многие тысячи – от простейших до некоторых пород индеек). Наконец, так называемые апомиктические растения, образующие семена из нормальных, диплоидных клеток с двойным набором хромосом, хотя бы одуванчики, каждую весну золотым потопом заливающие наши газоны. Или же растения в принципе способные к рекомбинации, но размножаемые черенками клубнями, отводками (картофель бананы и т. д. и т. д.). Кроссинговер, казалось бы, шире распространен, чем половой процесс, но, например, самцы дрозофил к нему не способны.
Отсюда делаем вывод: наличие генетической рекомбинации – не безусловный критерий живого. Это не условие жизни, а ее завоевание, не аксиома, а теорема (правда, одна из самых основных).
Мой коллега Алексей Владимирович Яблоков, ознакомившись с рукописью, предложил в качестве пятой аксиомы так называемое «давление жизни», геометрическую прогрессию размножения. Рассмотрим этот вопрос.
Еще Дарвин указывал, что такой важный фактор эволюции, как борьба за существование, вытекает из «быстрой прогрессии, в которой все органические существа стремятся размножиться». Здесь тот же принцип, какой положил в основу своего гонорара легендарный изобретатель шахмат: на первую клетку доски положить одно зернышко пшеницы, на вторую 2, на третью 4 и т. д., вплоть до 264 на последней клетке. Математики назвали такой рост геометрической прогрессией. Известно, что в таком случае прирост идет лавинообразно, на нем и основан рост делящихся атомов урана в ядерной бомбе. Таких примеров биологи приводили много. Дарвин писал, что «слон плодится медленнее всех известных животных», но за 740–750 лет потомство одной пары составило бы около 19 миллионов особей. Если же мы возьмем организмы с более быстрым темпом размножения, величины получаются буквально астрономические. Многократно упоминавшаяся нами плодовая мушка дрозофила за один год дала бы столько потомства, что оно покрыло бы землю слоем в миллион миль (!) толщиной, пара воробьев за десять лет расплодилась бы до 257 716 983 636 особей. Каждый из любителей математики, имея под рукой данные о плодовитости и продолжительности жизни каких‑либо организмов, на досуге может получить цифры не менее поразительные. Особенно если расчет относится к бактериям, которые в благоприятных условиях делятся, удваивают свою биомассу два раза в час.
Можно сказать, что это все теоретические выкладки, ничего подобного в природе не бывает. Не совсем так: если вид попадает в благоприятные условия, не ограничивается пищевыми ресурсами и не имеет врагов, численность его возрастает фантастически. Так бурно разросся упоминавшийся Дарвином чертополох в Аргентине, так размножались европейские кролики и американские кактусы‑опунции в Австралии и американская тля филлоксера на европейских виноградных лозах. Микроорганизмы, животные и растения, оставившие после себя залежи руд, целые пласты известняковых пород, все запасы угля и нефти, попадая в благоприятные условия, перестраивают облик всей нашей планеты.
Жизнь как бы использует любой ресурс, любую возможность для размножения. Это и есть «давление жизни». Но даже если численность организмов какого‑либо вида остается стабильной, потенциал его размножения – мощный резерв, поставляющий материал отбору.
Аксиома это или нет? Все же – нет, это следствие, вытекающее автоматически из принципа матричного воспроизведения. Это тоже не аксиома, а теорема, и тоже одна из главных, на которой основана вся современная биология.
И тем не менее я не могу безапелляционно утверждать что из массы положений, сформулированных биологами за последние 150 лет, я избрал в качестве аксиом именно те, которые этого названия заслуживают. И что их именно столько, сколько нужно для объяснения жизненных явлений – ни одной больше и ни одной меньше. Иными словами, отвечает ли эта система двум требованиям – полноте и независимости, как говорят математики.
Но, как я уже писал в предисловии, эта книга не учебник, а приглашение к раздумьям.
Впрочем, мои попытки вывести из аксиом многие эмпирические закономерности оказывались удачными. Так получилось с модусами эволюции или биогенетическим законом Мюллера – Геккеля, законом Долло, гласящим о необратимости эволюционных изменений, и целым рядом других. Однако чтобы представить хотя бы основные достижения биологии в аксиоматизированном виде, потребовалась бы книга раз в десять объемистей, чем эта. Предстоит работа не для одного человека и не на один год. Быть может, я продолжу эту тему дальше. Скажу более, именно этим я сейчас по мере своих возможностей и занимаюсь.
А пока, если эти очерки хотя бы выработают у читателя подобие иммунитета к высокопарным утверждениям о качественных отличиях живого от неживого (без расшифровки, что же это за отличия!), – о несводимости биологических закономерностей к физическим (без формулировки самих биологических закономерностей!), автор сочтет свою задачу выполненной.

[1]
Сегодня в полдень пущена ракета,
Она летит куда скорее света
И долетит до цели в семь утра
Вчера.
В этом шутливом английском стихотворении хорошо показан парадокс нарушения принципа причинности в неэйнштейновском мире.
[2]Первым подобный опыт проделал А. Лавуазье, поместивший в примитивный калориметр, охлаждавшийся льдом, морскую свинку. Измерив количество оттаявшей воды, он сопоставил его с количеством кислорода, потребленного свинкой, и пришел к выводу: жизнь – это медленное горение.
[3]ad infinitum – до бесконечности.
[4]См.: Эйген М., Винклер Р. Игра Жизни. М., Наука, 1979.
[5]Точнее, у финикийцев в начале слова был гортанный звук, отсутствующий в греческом языке; греки сочли возможным свой звук «А» передать финикийским символом α.
[6]В научной литературе этот процесс называют «рекогниция » – то же «узнавание », только по‑латыни.
[7]Медников Б. Дарвинизм в XX веке. М., Советская Россия, 1975.
[8]Синезеленые, строго говоря, не водоросли. Это одна из ветвей бактерий. Лучше назвать их цианофитами. Но старое название еще широко распространено, особенно в популярных изданиях.
[9]Впрочем, не все антрололаги разделяют его взгляд.
[10]Не могу не процитировать одно из писем: «Скорость света, объявленная беспричинно конечной скоростью распространения взаимодействий в мат. мире – это ничто иное, как неохристианство в материализме» (орфография и пунктуация подлинные). По‑видимому, автор этого письма искренне убежден, что если покрепче обругать, то и доводов не нужно. Впрочем, не он один…
[11]6 и 9 августа 1945 года американцы, как известно, сбросили на японские города Хиросиму и Нагасаки атомные бомбы, вызвав ничем не оправданные жертвы и разрушения.
[12]Есть, впрочем, исключения из этого правила. У некоторых растений (энотера) и животных (скорпионы) хромосомы, соединяясь друг с другом концами, образуют кольцо, передающееся по наследству как единое целое. Но у эукариотных организмов это вторичное исключение.
Photosynthesis and the Chloroplast
The activities of life require a continuous input of energy, which almost all organisms obtain by oxidizing complex organic molecules such as carbohydrates, fats, and proteins. Un- less constantly renewed, the existing levels of these molecules would be very quickly exhausted. The supply is continuously replenished by photosynthesis in the eukaryotic algae and higher plants and in the photosynthetic bacteria and blue-green algae.
Photosynthesis can be defined as a process in which sun- light is used as an energy source for converting electrons to a high-energy form. These electrons are then used to reduce or fix an electron acceptor, usually an inorganic substance, trans- forming it to a more complex organic molecule. There are thus two overall steps in photosynthesis: (1) transfer of light energy to electrons and (2) use of the electrons for reductions in cellu- lar synthesis. In the blue-green algae, eukaryotic algae, and higher plants, H2O is used as the source of the electrons in step of this process. The high-energy electrons are used to reduce another inorganic molecule, CO2, in step 2, fixing it into organic compounds. Oxygen, derived from the H2O used in step is released as an important by-product of the reaction. The bacteria capable of photosynthesis, the green and purple photosynthetic bacteria, use more highly reduced substances than H2O as an electron source and release molecules such as sulfur or hydrogen rather than oxygen as by-products.
Both steps of photosynthesis in eukaryotes take place inside the organelle specialized for this function, the chloroplast. Within chloroplasts, the enzymes and pigments carrying out step 1 (the absorption of light energy and the excitation of electrons to high energy levels) are concentrated in membranes; step 2 (CO2 fixation) is carried out by enzymes suspended in solution in the chloroplast interior. In prokaryotes, the reactions of photosynthesis are distributed between the plasma membrane and its derivatives, and the background substance of the cytoplasm.
Morphology and Major Molecular Components of Chloroplasts
Chloroplast Structure
The chloroplasts of algae and higher plant cells are easily seen under the light microscope as discoid or lens-shaped bodies with overall dimensions of about 5-10 µm. Usually chloroplasts are considerably larger than the mitochondria in the same plant cell and occur in smaller numbers. The number may vary from a single chloroplast, as found in Micromonas and several other green algae, to several hundred in the cells of most higher plants.
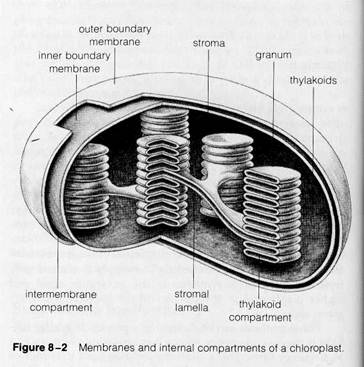
The green pigments of chloroplasts can be seen under the light microscope to be concentrated in disclike substructures, the grana, in most eukaryotic plants. Grana are suspended in a colorless background substance in the chloroplast interior. This pigment distribution is maintained by a system of membranes and compartments visible in chloroplasts under the electron microscope (Figs. 8-1 and 8-2). Two continuous boundary membranes, roughly analogous to the inner and outer boundary membranes of mitochondria, separate the chloroplast interior from the surrounding cytoplasm. The region between these membranes, the intermembrane compartment, is usually difficult to trace in electron micrographs (as in Fig. 8-1) because of extensive folding of the inner boundary membrane. The two boundary membranes enclose an interior compartment, the stroma, equivalent in location to the mitochondrial matrix. Within the stroma is a complex system of membranous sacs that forms the grana and their connections.
|

|
|
The individual unit of a granum is a flattened sac or vesicle, the thylakoid (from thylakos, meaning "sack" or "pouch"), consisting of a single, continuous membrane enclosing an interior thylakoid compartment. A granum is formed from a closely fused pile of these individual sacs, set one on top of another much like a stack of coins. Chloroplasts may contain from 40 to 60 grana, each formed from stacks of 2 or 3 to as many as 100 individual thylakoids.
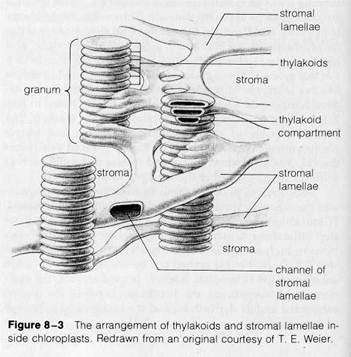
Numerous membranous connections can be seen between the thylakoids of adjacent grana (circled in Fig. 8-1). These connections, called stromal lamellae, enclose a channel that is continuous with the thylakoid compartments of adjacent grana (diagrammed in Figs. 8-2 and 8-3). The stromal lamel-
' lae may connect the thylakoid compartments into a single,
I continuous internal region inside chloroplasts.
Stacking and fusion of thylakoids into grana is typical of most chloroplasts of the green algae and higher plants. Elon-
I gated, single thylakoids appear along with grana in a few
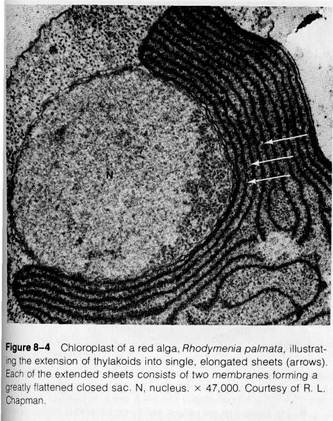
I chloroplast types in higher plants, most notably in the bundle sheath cells of some plants with the C4 photosynthetic pathway (see pp. 188-190). Unstacked thylakoids occur exclusively in nongreen algae. In the chloroplasts of red algae, thylakoids are single and greatly elongated (Fig. 8-4). In other nongreen algae, elongated thylakoids occur in multiple layers without fusion of adjacent membranes (Fig. 8-5 shows a chloroplast of this type from a brown alga). Despite these variations, the
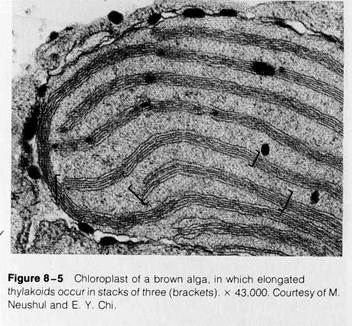
basic structure of thylakoids is uniform. Each thylakoid consists of a single, continuous membrane enclosing a flattened inner compartment that is completely separated from the surrounding stroma by the thylakoid membrane.
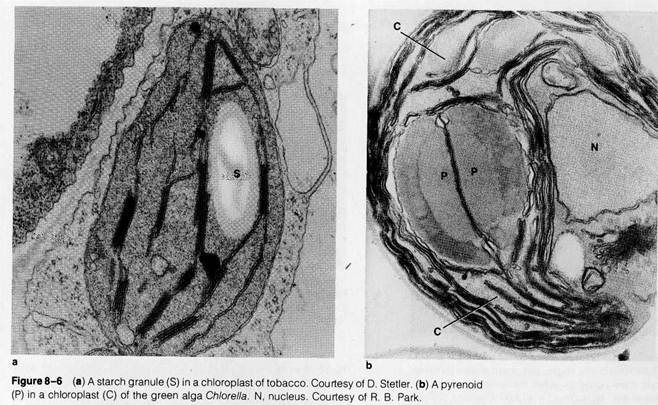
The chloroplast stroma may contain a variety of inclusions suspended in the regions surrounding the thylakoid and stromal lamellae membranes. Most conspicuous of these are starch granules (Fig. 8-6), found in chloroplasts after a period of active photosynthesis in the light. The stroma also frequently contains dense, spherical particles called osmiophilic granules or droplets (visible in Fig. 8-1), probably representing aggregates of lipids suspended in the stromal solution. In algae, large bodies called pyrenoids appear in the stroma (see Fig. 8-6b). Because these bodies are frequently surrounded by a layer of starch granules, they have traditionally been regarded as functioning in carbohydrate synthesis. This conclusion is supported by the work of R. H. Holdsworth (1971), who analyzed isolated pyrenoids isolated from a green alga. Holdsworth found an enzyme active in the initial step in carbohydrate synthesis (ribulose diphosphate carboxylase) to make up a large part of the mass of the pyrenoids.
The chloroplast stroma, like the mitochondrial matrix, contains DNA, ribosomes, and all of the biochemical factors required for DNA replication, RNA transcription, and protein synthesis. Of the visible elements of this system, the DNA appears at scattered locations in the stroma as faintly visible threads (see Fig. 12-43). The ribosomes, conspicuously smaller than the ribosomes of the surrounding cytoplasm, are distributed throughout the stroma (see Fig. 13-34; the functions of the chloroplast DNA and ribosomes in replication, transcription, and protein synthesis are discussed at length in Supplements 12-2 and 13-1).
|
|
|
Figure 8-8The chlorophylls. The structure of chlorophyll a is shown. Chlorophyll b has a —CHO group substituted for the —CH3 at the 3-carbon; chlorophyll d has a —CHO substituted for —CH—CH2 at the 2-carbon; protochlorophyll has double bonds instead of —H at the 7- and 8-carbons. The ring structure of the chlorophylls is similar to the prosthetic groups of the cytochromes and hemoglobin except for the additional segment shown (shaded portion).
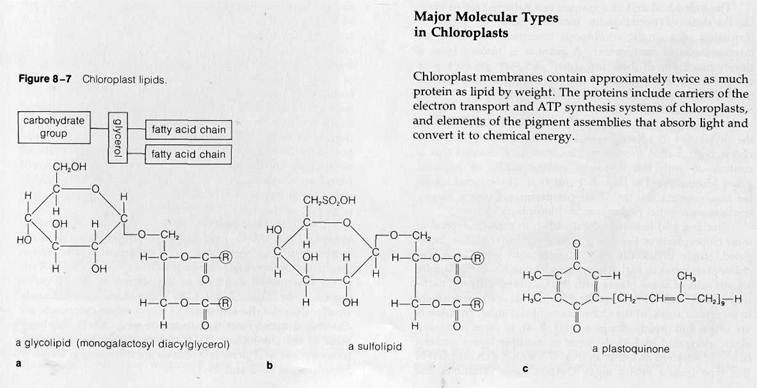
Glycolipids constitute about 45% of the lipid fraction. These lipids are built up from a glycerol backbone, combined with two fatty acid chains and a mono- or disaccharide carbohydrate group (Fig. 8-7). Some of the glycolipids, about 4% of the total lipid fraction, contain sulfur atoms as a part of their carbohydrate groups. One of these unusual glycolipids, termed sulfolipids, is shown in Fig. 8-7. Phospholipids, at about 10% of the total, form a much smaller fraction of the lipid content than they do in mitochondria. Chloroplast quinones, including plastoquinones (Fig. 8—7c) and vitamin K, make up about 6% of the total chloroplast lipids; some of these molecules function in photosynthetic electron transport. Sterols are present in chloroplast membranes at concentrations less than 1%. The remaining lipids of chloroplasts consist primarily of the photosynthetic pigments, the chlorophylls and carotenoids, which make up 28 to 30% of the total.
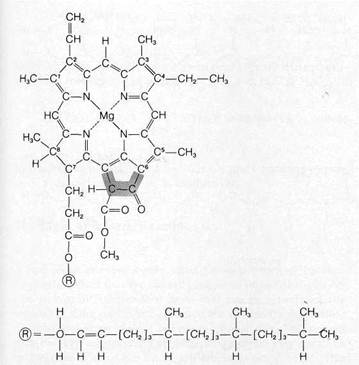
The predominant lipid pigments of chloroplasts are the chlorophylls, which absorb red and blue wavelengths most strongly and transmit green light. The various chlorophylls have in common a porphyrin ring to which a long phytol side chain (an alcohol) is attached (Fig. 8-8). The porphyrin ring of chlorophyll is similar to the prosthetic groups of the cytochromes and hemoglobin except for the presence of an extra
subunit (shaded in Fig. 8-8) linked at one side. A magnesium atom is bound at the center of the porphyrin ring in the chlorophylls. At least three major types, chlorophyll a, b, and c, which differ only in the side groups bound to the porphyrin ring, occur in different eukaryotic organisms.
All photosynthetic eukaryotes utilize chlorophyll a as their primary photosynthetic pigment. In the green algae and higher plants, chlorophyll b occurs as a second major photosynthetic pigment. Chlorophyll c occurs with chlorophyll a in the brown algae, diatoms, and dinoflagellates. Chlorophyll a is also found as the sole chlorophyll pigment in the blue-green algae. These chlorophyll pigments occur in combination with specific proteins, held in place in the amino acid chain of the protein by hydrophobic, noncovalent bonds (see, for example, Markwell, Thomber, and Boggs, 1979).
The wavelengths of light absorbed by the different chlorophylls vary depending on their state of binding within chloroplast membranes. Purified chlorophyll a, for example, absorbs light most strongly at wavelengths of 420 and 675 nm ^in acetone solutions. Within membranes, chlorophyll a may show other absorption maxima, at 660, 670, 678, 685, or 700 nm, along with the 420- and 675-nm maxima. These changes in absorption are due to interactions between the chlorophylls and between the chlorophylls and their binding proteins, and not to chemical changes in the chlorophyll molecules. The differently absorbing forms of chlorophyll a, for example, are chemically identical.
The carotenoids (Fig. 8-9) are also found in all photosynthetic organisms. These lipid-soluble pigments absorb light at blue wavelengths, from 400 to 550 nm, and transmit yellow and orange. All of the carotenoids are based on a long carbon chain containing 40 carbon atoms, linked together by double and single bonds. Various substitutions in the side groups attached to the 40-carbon backbone give rise to the different carotenoid pigments. Two types of carotenoids occur in both eukaryotes and prokaryotes. The carotenes are pure hydrocarbons; most abundant among these in higher plants is fi-carotene (Fig. 8-9a). The second type of carotenoid, the carotenols, contains hydroxyl (—OH) groups at both ends of the carbon chain. The yellow pigment xanthophyll (Fig. 8-9b) is the predominant carotenol in higher plants.
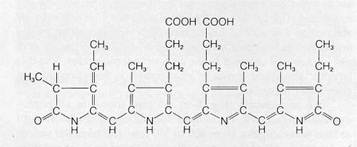
A final group of pigments is found in addition to chlorophyll a in the blue-green and red algae. These pigments, called phycobilins, contain a light-absorbing molecular structure resembling the porphyrin ring of hemoglobin except that the complex occurs in an extended, linear form (Fig. 8-10). Two kinds of phycobilins, the phycocyanins andphycoerythrins, occur in both the blue-green and red algae. In both groups, the phy-cobilin pigments occur in combination with proteins in complexes known as phycobiliproteins.
The carotenoids and phycobilins function as accessory pigments that take up light at wavelengths not directly absorbed by the chlorophylls. The light energy absorbed by these pigments and by chlorophylls b or c is eventually transferred to chlorophyll a, which is the photosynthetic pigment immediately involved in transforming light into chemical energy. The
■
8 Photosynthesis and the Chloroplast 173

|
Figure 8-9The carotenoids (3-carotene (a) and xantho-phyll (b).
|
Figure 8-10The light-absorbing structure of a phycocyanin.
net effect of the entire complement of photosynthetic pigments is to greatly broaden the spectrum of wavelengths usable as energy sources for photosynthesis.
Reactions of Photosynthesis The Overall Reaction
The basic elements of the generalized photosynthetic reaction synthesizing glucose in eukaryotes were worked out in the early nineteenth century:
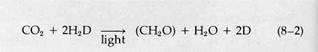
|
Subsequent research, extending into this century, established that some bacteria carry out photosynthesis using electron and H+ donors other than H2O. Many substrates, such as H2S, thiosulfate, alcohols, and hydrogen itself, can be used. This led C. B. Van Niel in 1935 to propose a more basic expression for the reactions of photosynthesis:
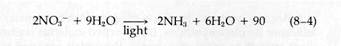
|
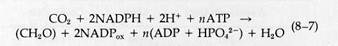
|
In this chapter, we will be concerned primarily with the photo-synthetic mechanism in which A is CO2 and H2D is H2O:
Reduced NADP is written as NADPH to keep track of the H+ ions removed from water. For the dark reactions,
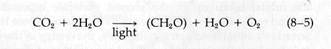
|
In the higher plants, six of the (CH2O) units combine to produce a six-carbon sugar such as glucose.
| <== предыдущая лекция | | | следующая лекция ==> |
| Oxidation of Pyruvic Acid to Two-Carbon Segments. | | | The Light and Dark Reactions |