Белки и ферменты, участвующие в репликации ДНК
В репликации ДНК принимают участие многочисленные белки и ферменты. Рассмотрим их основных представителей. ДНК-полимеразы ДНК-полимеразы осуществляют синтез ДНК. Субстратом для этих ферментов являются дНТФ: дАТФ, дГТФ, дЦТФ и дТТФ. Урав- нение реакции, катализируемой ДНК-полимеразами, в общем виде выглядит так:
ДНК-полимеразы последовательно добавляют нуклеотиды к3’ - концу полинуклеотидной цепи: Синтез новой цепи происходит в направлении от 5’-конца к 3’-концу. ДНК-полимераза может только наращивать цепь ДНК, начать же синтез ДНК с нуля она не в состоянии, для начала ее работы требуется затравка. В качестве затравки может выступать фрагмент ДНК или РНК (рис. 2.1). ДНК-полимераза способна удлинять цепь 23 также только в присутствии цепи, играющей роль матрицы (рис. 2.1). Нуклеотиды присоединяются к затравке в соответствии с принципом комплементарности, напротив аденина всегда будет встраивается тимин, а напротив гуанина – цитозин (рис. 2.1).
ДНК-полимеразы способны копировать любую цепь ДНК, т.е. они не специфичны к последовательности нуклеотидов матрицы. Скорость ДНК-полимеразной реакции колеблется в пределах от 50 нуклеотидов в секунду у эукариот до 500 нуклеотидов в секунду у прокариот. Главной задачей ДНК-полимераз является снятие точной копии с матрицы, в связи с этим они проверяют комплементарность каждого нуклеотида дважды: перед включением его в состав растущей цепи и перед тем как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется, если последний нуклеотид комплементарен матрице. Если включен некомплементарный нуклеотид, то он удаляется за счет 3 -> 5’-экзонуклеазной активности ДНК-полимеразы, и только после удаления некомплементарного нуклеотида ДНК-полимераза продолжит наращивать цепь ДНК (рис.2.2).
Рис.2.2. ДНК-полимераза удаляет некомплементарный нуклеотид и затем продолжает синтез ДНК.
Праймаза Праймаза катализирует матричный синтез короткой РНК- затравки (праймера) в направлении 5’→ 3’. Праймер использует ДНК-полимераза для инициации синтеза ДНК (рис. 2.3).
Рис. 2.3. Праймаза синтезирует РНК-затравку, которую ДНК-полимераза использует для синтеза ДНК. ДНК-лигаза соединяет 5’-фосфатную и 3’-гидроксильную группы соседних нуклеотидов, в результате образуется фосфодиэфирная связь, ликвидирующая разрыв (рис. 2.4).
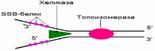
ДНК-хеликаза осуществляет расплетение двойной спирали ДНК, используя энергию гидролиза АТФ. В результате ее действия возникает «вилка» (Y), состоящая из двуцепочечного участка ДНК и двух одноцепочечных ветвей (рис. 2.5). Белки, связывающиеся с одноцепочечной ДНК (SSB-белки), обладают большим сродством к одноцепочечной ДНК, препятствуют образованию двойной спирали (рис. 2.5). Топоизомеразы снимают напряжения, возникающее в результате расплетения двойной спирали в ДНК, за счет разрыва и последующего воссоединения цепи ДНК (рис. 2.5).
Рис. 2.5. Роль ДНК-хеликазы, SSB-белков, топоизомеразы
|