ANNOTATED
Рис 1.4. Динаміка вирощування та реалізації птиці в Україні. Надалі прогнозується збільшення виробництва м’яса птиці та яєць. За дев’ять місяців 2013 року вирощено 1086,9 тис. т м’яса птиці в живій вазі, що на 12,9% перевищує рівень 2012 року. Виробництво яєць становило 16,6 млн шт. на 2013 рік, що на 2,4% більше минулорічного. Очікується, що збільшення вітчизняного виробництва м’яса птиці позитивно позначиться на зовнішньоторговельній діяльності, тобто зменшенні його імпорту та зростанні експорту. За інформацією Міністерства доходів і зборів за січень-жовтень 2013-го було експортовано 118,2 тис. т м’яса птиці і субпродуктів, що удвічі більше, порівняно з аналогічним періодом 2012 року. У грошовому виразі експорт цієї продукції зріс на 82,6%, до 192,5 млн дол. США. У той же час, імпорт м’яса птиці і субпродуктів за цей період скоротився на 37,4%, до 60,9 тис. т, а в грошовому еквіваленті - на 38,6%, до 82,7 млн дол. США.
Рис 1.5. Динаміка виробництва яєць в Україні. При зовнішніх продажах України й надалі віддаватиметься пріоритет м’ясу птиці. Птахівництво розвивається більш інтенсивно. До того ж, в січні 2013 року Україна отримала дозвіл на продаж птиці та яєць У ЄС. Це дає великі шанси виходу на нові ринки, хоча суттєві тарифні бар’єри в ЄС будуть стримувати цей процес. Розвиток галузі сприятиме збільшенню споживання м’яса та м’ясопродукції. Якщо середньорічне споживання м’яса на душу населення збільшилося із 46,6 кг у 2011 році до 54,4 кг у 2012-му, то прогнозується, що в 2013 році воно становитиме 55,3 кг. При цьому річне споживання м’яса птиці зросте із 23,8 кг на одну особу до 24,5 кг. Основний вплив на розвиток ринку м’яса в Україні матимуть ціни на корми, податковий режим для виробників. У той же час, роль прямої державної підтримки суттєво знижуватиметься, враховуючи незначні фінансові можливості бюджету наступного року [50]. 1.2. Характеристика пташиного посліду Пташиний послід – це продукт обміну речовин в організмі птиці колоїдної консистенції, сіро-зеленого кольору, комкувато-пористої структури, що виділяється з організму птиці у вигляді суміші сечі та калу. Вихід посліду залежить від виду птиці, її віку та способу утримання. Усереднена норма виходу посліду з урахуванням усушки до 65-70% становить на одну голову дорослої птиці 62 кг на рік. При зберігання у чистому вигляді послід швидко злежується та випускає неприємний запах, зумовлений виділенням продуктів розкладу органічних речовин. Більша частина посліду складається з часток розміром від 0,1 до 1 мм, а близько 30% його маси становлять частки розміром менше 0,1 мм. Дрібні фракції містять більше органічної речовини (до 75%), ніж крупні (до 25%). У посліді містяться включення неперетравленої їжі, перо птиці, пісок та ін. Усі поживні речовини у пташиному посліді знаходяться у сполуках, які здатні засвоювати рослини. Послід також багатий на мікроелементи: у 100 г сухої речовини міститься мангану15-38 мг, цинку – 12-39, кобальту – 1-1,3, міді – 0,5, заліза – 367-900 мг. Значна частина елементів живлення в посліді (азоту близько 50%, фосфору – 40% та калію – 60%) знаходяться у водорозчинній формі. Додавання води у послід значно знижує його цінність як добрива. Хімічний склад пташиного посліду яєчних курей промислового стада у% на суху речовину: вологість – 71%, N – 1,24, P2O5 – 0,57, K2O – 0,51. При сухому способі видалення посліду його вологість складає 55-60%. Зазвичай вода з напувалок в тій чи іншій мірі потрапляє на настили та зволожує послід, тому абсолютно сухого способу видалення посліду з кліткових батарей поки не існує [19, 20]. Непідготовлений до використання гній та послід містять яйця та личинки гельмінтів, патогенну мікрофлору, життєздатне насіння бур'янистих рослин. Можлива наявність збудників інвазійних та інфекційних хвороб. Екологічна небезпека гною та посліду також багато в чому визначається значною концентрацією в них токсичних сполук типу аміаку, сірководню, меркаптанів, фенолу, крезолу та ін. Особливу небезпеку становлять леткі азотовмісні сполуки. Курячий послід офіційно визнано хімічним та бактеріологічним токсичним компонентом третього класу небезпечності [19]. 1.3. Методи та способи переробки пташиного посліду З переходом птахівництва на промислову основу й появою великих птахівницьких господарств, поголів'я птиці в яких становить десятки й сотні тисяч голів, галузь зіштовхнулася із проблемою використання великої кількості пташиного посліду [8]. Методи та способи переробки пташиного посліду залежать від властивостей і складу початкової сировини, а також від цілей переробки та виду кінцевих продуктів [19, 20]. При виборі методів і способів переробки пташиного посліду виходять з науково обґрунтованої системи поглядів, а саме: - пташиний послід має енергетичний потенціал, обумовлений вмістом органічних речовин, здатних при їх мінералізації віддавати значну кількість енергії; - пташиний послід є невід'ємною ланкою природного ланцюга "ґрунт – рослина – тварина (птахи) – гній (послід) – ґрунт", і повинен використовуватися в цьому ланцюзі з найбільшим ефектом; - будучи залишком переробки продукції фотосинтезу рослин птахами, пташиний послід містить велику кількість органічних речовин, біогенних елементів, мікроелементів, ферментів і вітамінів; - наявність органічних речовин, робить пташиний послід ідеальною їжею для ґрунтової мікрофлори, життєдіяльність якої забезпечує нормальний перебіг біохімічних процесів у ґрунті, сприяє утворенню гумусу і збільшенню родючості ґрунту; - біологічні особливості пташиного посліду унеможливлюють його пряме застосування в якості органічного добрива, через невідповідність необробленого посліду санітарно-ветеринарним вимогам; - кінцеві продукти переробки пташиного посліду не повинні містити умовно патогенну мікрофлору, яйця гельмінтів і їх застосування не повинно викликати захворювання людей і тварин, накопичення шкідливих речовин у ґрунті; - вміст біогенних речовин у пташиному посліді дозволяє суттєво знизити застосування мінеральних добрив, для виробництва котрих необхідні значні витрати сировинних та енергетичних ресурсів; - переробка посліду є важливим економічним, господарським та природоохоронним завданням, розгляд якого повинен відбуватись з позиції системного підходу [19, 21]. Виходячи з вищесказаного, переробка пташиного посліду може проводитись в наступних напрямах: - отримання органічного добрива; - палива; - з метою знищення; - комбіновано [21]. Для цього можуть застосовуватись різні методи переробки гною, які можна розділити на фізичні, хімічні і біологічні. До фізичних методів відносяться: механічні, теплові та електричні. Механічні методи включають ряд способів обробки рідкого пташиного посліду, такі як відстоювання, центрифугування, фільтрування. Представлені способи спрямовані на зневоднення посліду і розділення його на фракції. Способи розділення пташиного посліду на фракції і зневоднення рідкого посліду умовно можна розділити на природні і механічні. Природне розділення рідкого посліду відбувається в горизонтальних і вертикальних відстійниках під дією гравітаційного поля землі. Ці пристрої прості, але природне відстоювання посліду не відбувається, якщо вологість його менше 90%. Продуктивність відстійників невелика. Крім того, вони громіздкі і дорогі. Проте, ці пристрої знаходять застосування в технологічних лініях розділення тваринницьких стоків. Механічне розділення рідкого посліду на фракції являється основним видом сепарації стоків на великих комплексах. Розділення відбувається за допомогою фільтруючих та осаджуючих машин та апаратів, загальним недоліком яких є висока вологість одержаної твердої фракції. Серед фільтруючих машин та апаратів найбільше розповсюдження отримали прес-фільтри та вібросита. Механічні методи можна розглядати як допоміжні, оскільки вони не вирішують проблеми знезараження при переробці пташиного посліду і є досить енергоємними. Отримані рідка та тверда фракції вимагають подальшої обробки біологічними, хімічними або тепловими способами. До теплових способів переробки посліду відносяться сушка, обробка гострою парою і випарювання. Сушку можна здійснювати як низькотемпературним, так і високотемпературним способом. Низькотемпературна сушка здійснюється за природних умов та дозволяє призупинити бродіння посліду. При застосуванні цього способу значно знижується вологість посліду без залучення значних витрат енергії, зменшується неприємний запах, але немає гарантії повного знезараження. Також в процесі сушки відбуваються істотні втрати цінних компонентів посліду. В останні роки отримав поширення спосіб сушки посліду у спеціальних агрегатах під дією високих температур. Установки для сушіння посліду забезпечують отримання продукту, що відповідає санітарно-ветеринарним вимогам, зручного для транспортування і внесення в ґрунт. Проте, висока вартість високотемпературної сушки обумовлена ціною на паливно-мастильні матеріали і високими первинними витратами, що не дозволяють широко застосовувати цей спосіб переробки. Обробка гострою парою спрямована на повне знезараження посліду. Цей спосіб енергоємний і послід на виході має більш високу вологість, що пов'язано з розрідженням його парою. Застосування цього способу можливо у відносно вузьких виробничих умовах і у випадку виникнення епізоотії. Спосіб термічної обробки посліду в автоклавах застосовується для знезараження рідкого пташиного посліду, але вимагає значних затрат енергії. До групи електричних методів відносяться обробка струмом надвисокої частоти, електроосмос, електрохімічна коагуляція, електрофлотація, обробка ультрафіолетовими променями. Ці способи не отримали широкого поширення через високу вартість устаткування, значні витрати електроенергії та підвищені вимоги, що пред'являються до персоналу. Одним з видів обробки рідкого пташиного посліду є хімічні методи. Вони запобігають забрудненню навколишнього середовища та виділенню неприємних запахів в довкілля. Крім того, деякі хімічні реагенти знищують бактерії, у тому числі й ті, що викликають гнилісні аеробні процеси, що супроводжуються виділеннями шкідливих газів. Хімічні методи передбачають контактну обробку хімічними речовинами і застосовуються тільки для знезараження посліду. Загальним недоліком хімічних методів є те, що вони вимагають певних матеріальних та енергетичних витрат, а також хімічні речовини чинять певне навантаження на навколишнє середовище [20, 21, 10]. Перспективним напрямом переробки пташиного посліду вважаються біологічні методи. Вони засновані на біохімічній деструкції та мінералізації органічних речовин мікроорганізмами і максимально наближені до природних процесів. При цьому за рахунок часткового розкладання органічної речовини відбувається зменшення вмісту в посліді джерел забруднення, які переходять в нерозчинну або газоподібну форму. Біологічні способи підрозділяють на природні і штучні. Природні ґрунтуються на біохімічних процесах, що протікають в природних умовах - біологічних ставках, лагунах, ґрунті та компості. Штучні - засновані на біохімічних процесах, що протікають у штучно створених умовах, – в аеротенках, окислювальних каналах, метантенках, анаеробних фільтрах та інших спорудах. Залежно від того, які групи мікроорганізмів беруть участь в процесі, розрізняють аеробні і анаеробні способи біологічної обробки. Компостування посліду відноситься до аеробних біологічних процесів розкладання. Знезараження посліду відбувається при його розкладанні та нагріванні (до 60 °С) при наявності термофільних мікроорганізмів. Основними недоліками цього способу є обмеженість застосування за кліматичними зонами країни, тривалість процесу, значні втрати поживних речовин та необхідність великої кількості наповнювача. Спосіб обробки рідкого пташиного посліду у біологічних ставках має відносно низьку вартість переробки. Проте цей спосіб тривалий в часі, вимагає відчуження значних виробничих площ. Також існує небезпека промерзання, підвищеного неприємного запаху, а також забруднення ґрунтових вод. Для інтенсифікації процесу розкладу та осадження органічної маси у пташиному посліді застосовують аеробну обробку. Вона відбувається шляхом аерації за допомогою нагнітання повітря від компресора, або за допомогою механічних аераторів, які за рахунок інтенсивного перемішування рідкого посліду забезпечують його контакт з атмосферним повітрям, що створює сприятливі умови для життєдіяльності аеробних бактерій. Але цей спосіб має ряд недоліків: значні втрати азоту, складність та високу вартість споруди, високі експлуатаційні витрати, а також великі витрати енергії на аерацію та безперервне перемішування. Анаеробне зброджування вважається перспективним способом. В результаті процесу зброджування розпаду підлягають органічні речовини, що містяться в посліді, з утворенням газоподібних продуктів у вигляді суміші метану і вуглекислого газу (біогазу). При цьому він частково або повністю знезаражується, дегельмінтизується, дезодорується та девіталізується. Для анаеробного зброджування застосовують різні біореактори: метантенки і анаеробні фільтри. Для забезпечення процесу потрібна підтримка температурного режиму, забезпечити яку можна шляхом використання біогазу, що виділився. Анаеробне зброджування покращує властивості посліду, як добрива, в результаті мінералізації біогенних речовин, практично без втрат їх у навколишнє середовище. Цей спосіб дозволяє більш високі норми навантаження, ніж можливі при аеробній обробці, не вимагає застосування хімічних реагентів для розкладання органічної речовини; зменшує водоутримуючі властивості посліду. Анаеробне зброджування – єдиний спосіб з вище розглянутих, що дозволяє не лише покривати витрати енергії на ведення процесу, але і отримувати надмірну її кількість. Енергія, що одержується у вигляді біогазу, зручна для користувача, оскільки її можна перетворити в теплову, електричну і механічну. Але цей метод практично не дає зменшення об’єму та вологості обробленого посліду [21, 10, 5]. Розглянувши основні методи і способи переробки пташиного посліду, можна зробити наступний висновок: найбільш відповідає пред'явленим вимогам спосіб анаеробного зброджування, як найбільш наближений до природних умов і який дозволяє отримати високоякісне органічне добриво і додаткове джерело енергії у вигляді біогазу. 1.4. Анаеробна ферментація пташиного посліду Для посліду характерним є значний вміст сечової кислоти та неперетравлених білків, які є джерелом N. Відношення С/N є меншим за оптимальне. За таких умов може утворюватися аміак, що за певних концентрацій призводить до інгібування процесу. За своїми фізичними та хімічними властивостями послід не дуже добре підходить для анаеробного зброджування, проте ряд дослідників повідомили про успішну роботу систем виробництва біогазу з пташиного посліду. Більшість з них були розроблені для роботи у мезофільному режимі (близько 35 °C) [28]. Хасан і співавт.для оптимального виробництва біогазу рекомендують використовувати послід з кількістю сухих речовин 4,6-7,6%, за температури 35 °С. Також повідомляється, що додавання речовин, що містять легко доступний вуглець і збільшення кількості посівного матеріалу інтенсифікує процес [28]. Ґрунтуючись на аналізі доповідей про анаеробне зброджування і виробництво біогазу з пташиного посліду, Сміт рекомендує використовувати співвідношення C:N на рівні 16:1, 40-денний час утримання, швидкість завантаження 2,4 кг сухих органічних речовин в день на м3 біогазової установки, об’єм реактора – 3,86 м3 на 1000 кг живої ваги тварин [28]. Інгібуючий ефект аміачного азоту був показаний рядом авторів (наприклад, P.L. McCarty і R.E. McKinney, P.N. Hobson і B.G. Shaw, J.W. Koster і G. Lettinga). Проте неодноразово показано, що метаногени можуть адаптуватися до підвищення концентрації амонійного азоту. Ряд дослідників повідомили про успішну роботу систем виробництва біогазу з пташиного посліду. Більшість з них були розроблені для роботи у мезофільному режимі (близько 35 °C). Parkin і Miller (1983) виявили інгібування амонієм при більш низьких концентраціях в умовах, коли система знаходилась при більших температурах. Далі наведено огляд досліджень у термофільному режимі [40]. Huang і Shih (1981) вивчали потенціал біологічного виробництва метану з курячого гною. Процес проводили у термофільних умовах при концентрації сухих органічних речовин від 1 до 8% і періоді обороту реактора від 3 до 10 днів. Короткий період обороту реактора супроводжувався вищим виробництвом газу, але зниження СОР і ХСК зменшувалось. При концентрації 8% СОР виробництво газу впало. Концентрація СОР 6%, тривалість 4 дні, рН 7,5 і температура 50 ° С були визначені оптимальними. При даних умовах вихід газу становив 4,5 л / л день, а метану 3,2 л / л день або 267 л / кг СОР [32]. Steinsberger і Shih (1984) вивчали роботу біогазової установки, що складалася з двох горизонтальних реакторів з'єднаних у серію. Корисний об'єм першого реактора становив 8 м3, другого реактора - 16 м3. Температура у першому реакторі підтримувалась на рівні 50 ° С. У другому не підтримувалась і становила 30-35 ° С. При концентрації інфлюенту 3% СОР, час утримання субстрату 4 дні і швидкості завантаження 7,5 кг СОР / м3 день вихід газу становив 3,9 м3 / м3 день з вмістом метану 55%. Збільшення швидкості завантаження до 9,4 кг СОР / м3 день з 5-денним RT і концентрацією інфлюенту 4,5% СОР викликало початкове збільшення виходу біогазу протягом тижня перед стійким його зниженням. Dosoretz i Lamed (1987) досліджували метаногенез курячого посліду при термофільному режимі (55 ºC) після тривалих процедур акліматизації. Вміст СОР становив від 24 до 96 г/л, час утримання від 5 до 20 днів. Метан утворювався у кількості від 0,5 до 0,7 л · л-1 день-1. Водень у біогазі не був присутній. Концентрації ацетату (від 19 до 222 ммоль), інших жирних кислот, і амонійного азоту (від 800 до 2830 мг/л) були дуже високими. Дослідження міченого атому показали, що ацетат накопичувався в результаті інгібування його переходу в метан. Марченко (2009) проводив анаеробне бродіння курячого посліду при вологості субстрату 92% і гідравлічному часі утримання 10 діб. Температуру змінювали від 48 до 58 оС. Вихід біогазу збільшувався при підвищенні температури. При цьому спостерігалося зменшення його приросту. З врахуванням енергетичних витрат на нагрівання субстрату отримали оптимальну температуру - 54 оС. Результати отримані в лабораторний умовах були перевірені на промисловій установці. Продуктивність установки становила по добриву 3 м3 на добу, а по біогазу - 75 м3 на добу. Вихід газу відповідав кривій, отриманій у лабораторних умовах. Abouelenien і співавт. (2009) вивчали сухе анаеробне бродіння курячого гною. Курячий гній був змішаний з активним мулом у ваговому співвідношенні 1:1 і інкубували при 37 ° C, 55 ° C і 65 ° C. Коли виробництво газу зупинилось половину вмісту склянок замінювали свіжим гноєм. Ця процедура була повторена 9 разів протягом 418 днів. Хоча виробництво біогазу спостерігалося при всіх температурах, метан не вироблявся при 55 і 65 ° C. Хамоков і співавт. (2012) здійснювали оптимізацію режимів роботи установки для переробки пташиного посліду. Процес проводили при температурі 50 ◦C, 55 ◦C і 60 ◦C, вологості субстрату 85%, 90% і 95% та гідравлічному часі утримання 7, 10 і 13 днів. З математичної моделі були отримані оптимальні параметри процесу: температура 54 ◦C, вологість субстрату 90%, тривалість переробки 11 днів. Вихід біогазу при цих значеннях основних факторів становив 0,67 м3/кг СР. Інгібуючий ефект аміачного азоту був показаний рядом авторів(наприклад, Маккарті і Мак-Кінні, Хобсон і Шоу, Костер і Леттінга). Проте неодноразово показано, що метаногени можуть адаптуватися до підвищення концентрації амонійного азоту [31, 36, 38]. З. Печан і співавт. вивчали бродіння курячого посліду при мезофільних температурах. У середньому вміст сухих речовин становив від 11,3% до 14,1%, сухих органічних речовин від 7,8% до 9,7% і тривалість утримання від 27 до 58 діб. Шкідливого впливу на виробництво біогазу не спостерігалося, незважаючи на середні концентрації амонійного азоту в стічних водах від 4,07 до 5,85 г/л. Вихід біогазу становив від 0,239 до 0,370 л/г сухих органічних речовин, вміст метану в біогазі - від 59 до 67% [28]. Ф. Абаюлянін і співавт. повідомляють про зброджування посліду вологістю 75% у мезофільних умовах при 37 °C з використанням системи повторної партії культури [23]. 1.5. Можливості використання біогазу на птахофабриках Одним з шляхів раціональної утилізації пташиного посліду є його анаеробне збродження, яке забезпечує одержання біогазу і високоякісного біологічного добрива. Виходячи з того, що птахи погано засвоюють енергію рослинних кормів та те, що більше половини цієї енергії використовується непродуктивно – йде у послід, його можна розглядати не лише як цінну сировину для органічних добрив, але і як потужне відновлюване джерело енергії. Біогаз — це газ, що утворюється в результаті метанового бродіння біомаси. Розклад біомаси проходить під впливом бактерій класу метаногенів. Склад: 55%-75% метан, 25% - 45% вуглекислий газ, незначні домішки. Енергія, що утворюється при спалюванні біогазу, може досягати від 60 до 90% тієї, якою володіє вихідний субстрат. Біогаз, що утворюється при бродінні має теплоту згорання 5340-6230 ккал/м3 (6,21-7,24 кВт·год/м3) [14]. Вихід газу з 1 кг сухої речовини пташиного посліду складає 0,31-0,32 м3. З 1 т субстрату вологістю 85% можна отримати 47-94 м3 біогазу. Вміст метану становить 60%. На отриманому біогазі можуть працювати газоспалювальні прилади, що виробляють енергію, яка використовується для опалення, освітлення, постачання кормозаготівельних цехів, для роботи водонагрівачів, газових плит, інфрачервоних випромінювачів і двигунів внутрішнього згорання. Побутові газові пальники споживають 0,2 – 0,45 м3 біогазу за годину, а промислові – від 1 до 3 м3 біогазу за годину [3]. Для опалення житлових будинків та адміністративних будівель у зимовий період необхідно у середньому 0,3 кВт·год теплової енергії на 1 м2 житлової площі. Кількість тепла необхідна для опалення тваринницьких ферм залежить від їх конструктивних особливостей, необхідної температури, теплоізоляції, вологості повітря, кількості та розміру тварин. Для курчат становить близько 0,2 кВт/м2. Питомі витрати тепла на обігрів блока парників площею 1 га при піковій розрахунковій різниці температур -32ºС складає 0,359 кВт/м2. На висушування 1 тони зерна від 19 до 14% вологості необхідно 70-90 кВт·год теплової енергії (12-15 м3 біогазу), 1 тони кукурудзи з 30 до 14% вологості — 250-300 кВт·год (42-50 м3 біогазу), 1 тону соняшника від 13 до 8% вологості — 160-180 кВт·год тепла (27-30 м3 біогазу) [18]. Досвід показує, що біогаз економічно доцільно використовувати в газоелектрогенераторах, при цьому спалювання 1 м3 біогазу дозволяє виробляти від 1,6 до 2,3 кВт·год електроенергії. Ефективність такого використання біогазу підвищується за рахунок використання теплової енергії, що утворюється при охолодженні двигуна електрогенератора, для опалення реактора біогазової установки. Ефективність використання біогазу складає 55% для газових плит, 24% для двигунів внутрішнього згорання. Найбільш ефективний шлях використання біогазу – у якості комбінації тепла і енергії, при якому можна досягти 88% ефективності. Біогаз можна використовувати у якості палива для автомобільних двигунів, при цьому ефективність його у даному випадку залежить від вмісту метану і наявності домішок. На метані можуть працювати як карбюраторні, так і дизельні двигуни. Однак, так як біогаз є високооктановим паливом, більш ефективне його використання в дизельних двигунах. Для роботи двигунів необхідна велика кількість біогазу і установка на двигуни внутрішнього згорання додаткових приладів, які дозволяють їм працювати як на бензині, так і на метані. Для використання біогазу у якості палива для двигунів внутрішнього згорання необхідне попереднє очищення біогазу від води, сірководню і вуглекислоти [3].
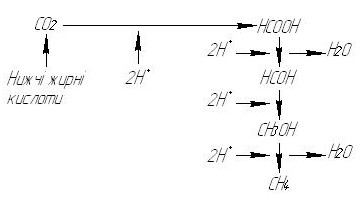
1.6. Біохімія метанового бродіння Процес метанового бродіння вивчався спочатку без зв'язку його з енергетикою. В результаті при вивченні механізму утворення метану дослідники прийшли до висновку про те, що хімізм утворення метану — це енергетичний процес, що забезпечує утворення АТФ, який використовується збудниками метанового бродіння. Оскільки до цього часу були відомі конкретні механізми енергетичних процесів, зокрема їх зв'язок з переносом водню і електронів, виникло припущення, що джерелом енергії при метановому бродінні є процес відновлення речовин воднем в метан, тобто перенос водню. Це відповідає загальнобіологічним уявленням про енергетичний обмін. Проте виникають питання, які речовини є акцепторами водню, які його джерела та шляхи переносу? Виявилось, що все це важко пояснити на основі тих уявлень, які склались спочатку відносно фізіології живлення метаноутворюючих бактерій, згідно з якими, “істинне” метанове бродіння — це перетворення нижчих жирних кислот і СО2 в метан. Причому з нижчих жирних кислот головну роль відіграє оцтова кислота, оскільки вона найбільш вірогідний продукт анаеробного окиснення органічних речовин. Однак просте відновлення жирних кислот дає відповідні вуглеводні. У такому випадку у метан може перетворитись лише мурашина кислота, але вона в процесі метаболізму утворюється у малій кількості. У свою чергу, з оцтової кислоти повинен утворюватись етан, однак цей вуглеводень майже не присутній у складі газоподібних продуктів метанового бродіння. Дослідники прийшли до висновку, що оцтова кислота у ході метанового бродіння піддається не відновленню, а розпаду, причому вуглекислий газ і метильна група, що утворюються своїми шляхами перетворюються в метан. Ці шляхи сходяться, так як відновлення метильної групи — це одна з останніх стадій хімізму відновлення вуглекислого газу в метан. У наш час вже не багато дослідників притримуються думки, що метаноутворюючі бактерії споживають тільки нижчі жирні кислоти і СО2. Проте хімізм утворення метану досі пов'язують з нижчими жирними кислотами, так що його не можна пояснити з точки зору енергетики. Згідно з існуючою думкою, утворення і відновлення СО2 йде за схемою:
Рис. 1.6. Схема утворення і відновлення СО2 при метановому бродінні.
При цьому всі проміжні продукти знаходяться у вигляді радикалів, пов'язаних з переносниками (тетрагідрофолієвою кислотою, коферментом, що містить вітамін В12, та ін.). У чистому вигляді вказані проміжні метаболіти (як це для спрощення показано на схемі) не виявляються. У реальності схеми відновлення СО2 в метан не може бути сумніву, хоча це не виключає інші можливості утворення метану, подібно тому, як H2S або NH3 утворюються не тільки у результаті редукції сульфатів і нітратів, але і при розкладі амінокислот або інших речовин. Протиріччя у наведеній схемі виникає у відношенні джерел активного водню. У наведеній схемі таких джерел знайти не можна. Нижчі жирні кислоти є кінцевими продуктами анаеробного метаболізму саме тому, що не можуть піддаватись подальшому окисненню, тобто бути джерелом активного водню. Робилися спроби показати, що відновлення СО2 проходить молекулярним воднем, що виділяється при окисненні білків, амінокислот, вуглеводів та інших речовин супутніми мікроорганізмами (вважаючи, що метаноутворюючі бактерії не здатні до розкладу цих речовин). При цьому передбачається, що метаноутворюючі бактерії, маючи здатність виробляти фермент гідрогеназу, використовують для відновлення СО2 водень ендогенного походження. Проте надзвичайно висока енергія розщеплення молекулярного водню, відома в хімії як енергія дисоціації водню, є непрямим свідоцтвом того, що такий механізм утворення активного водню в анаеробних умовах маловірогідний. Інша справа — вилучення атомарного водню з яких-небудь речовин, де він знаходиться в менш міцному хімічному зв'язку. У хімії таким прикладом є вилучення водню дією кислоти на метал. У момент виділення водень знаходиться в активній формі. У біологічній хімії аналогічну активацію водню органічних речовин здійснюють дегідрогенази. Біологічно активний водень — НАД·Н2 — це свого роду форма “in statu nascendi”, отримувана в хімії витісненням з кислот металами і використовувана на відновні реакції. Дегідрогенази тут виконують роль стабілізаторів, що перешкоджають швидкому сполученню атомарного водню в неактивну молекулярну форму. Продовжуючи цю аналогію, можна сказати, що у живому організмі, як і в хімії, такий шлях вилучення активного водню значно легше ніж, ніж дисоціація молекулярного водню за допомогою гідрогенази. Однак, приймаючи це, потрібно визнати, що метаноутворюючі бактерії, вилучаючи активний водень, окиснюють різноманітні органічні речовини, тобто здійснюють звичайне гетеротрофне живлення, причому споживання нижчих жирних кислот для них не обов'язкове. Відносно активації молекулярного водню метанутворюючими бактеріями, то експериментально це підтвердити не вдається. Крім того, це суперечить переконанню тих же авторів про те, що метанове бродіння може проходити на середовищі, що містить тільки нижчі жирні кислоти, коли виділення молекулярного водню взагалі неможливе. Всі протиріччя легко вирішуються, якщо уявити, що процес розкладу речовин з утворенням СО2 і активного водню, а потім утворення метану здійснюють одні й ті ж мікроорганізми, причому вихідними речовинами є не нижчі жирні кислоти, а білки, амінокислоти, вуглеводи та інші сполуки з високим енергетичним потенціалом. Тому слід визнати, що метанутворюючі бактерії споживають будь-які речовини. Для заперечення цього зараз вже майже немає підстав. При такому уявленні про хімізм утворення метану напрошується порівняння його з процесом відновлення сульфатів та нітратів. Про це ще в 1936р. згадував відомий американський дослідник метанового бродіння Г. Баркер, однак відносно джерел водню, необхідного для цього процесу, він притримувався іншої думки. Питання про вихідний стан СО2, що піддається відновленню, також потребує уточнення. По аналогії з редукцією сульфатів і нітратів, у процесі утворення метану відбувається перенос водню на кисень СО2. Однак, чи можливе акцептування водню молекулою вуглекислого газу, що знаходиться у недисоційованій формі? Швидше всього, для цього необхідні карбонат-йони, тобто вугільна кислота, що утворюється при хімічній взаємодії СО2 з водою: СО2 + Н2О → Н2СО3 → СО3+2 + 2Н+. Такому висновку, здавалося б, суперечать відомі дані, що СО2 слабко взаємодіє з водою і знаходиться у ній в основному просто у розчиненому вигляді. Однак так буває у чистому вигляді. У складному середовищі, де присутні різні речовини, у тому числі і мінеральні сполуки, певна кількість хімічно зв'язаного СО2, тобто вугільної кислоти, знаходиться у вигляді карбонатів. Вони дисоціюють, утворюючи карбонат-йони, які відновлюються в метан. По мірі їх відновлення, тобто зникнення з середовища, відбувається поповнення кількості зв'язаного СО2, відновлюється рівновага, і кількість карбонатів у середовищі стабільне і достатнє для утворення метану. Іншими словами, у процесі метанового бродіння у середовищі постійно підтримується певний рівень концентрації карбонатів, що забезпечує безперервне перетворення СО2 в метан. Слід врахувати, що в середовищі складного складу є і інші можливості зв'язування СО2, наприклад взаємодія з аміногрупами амінокислот і білків, з гідроксильними групами органічних сполук і т. д. Можливо, такі форми зв'язаного СО2, подібно карбонатам, дисоціюють і відновлюються в метан. Процес утворення метану із СО2 можна представити як процес редукції карбонатів:
Рис. 1.7. Утворення метану із СО2 представлене як процес редукції карбонатів.
Символом RH2 в схемі позначені будь-які речовини, що здатні окиснюватись (відщеплювати водень) в анаеробних умовах. Запропонована схема відрізняється від попередньої тим, що окиснюваним субстратом можуть бути будь-які речовини, і їх окиснення є одночасно джерелом СО2 і активного водню, за допомогою якого відбувається відновлення СО2 в метан. Все це здійснюють одні і ті ж мікроорганізми. Процес переносу водню на кисень вони використовують у якості джерела енергії. Відносно нижчих жирних кислот, зокрема оцтової, яка неминуче утворюється в анаеробному процесі, можна сказати, що метанутворюючі бактерії вилучають енергію до її утворення, а не з неї. Слід відзначити, що при розгляді хімізму утворення метану виникає питання не тільки про джерела активного водню. Враховуючи відсутність яких-небудь даних відносно джерел СО2, може здатися само собою зрозумілим, що його виділяється достатньо в процесі декарбоксилювання речовин, що споживаються. Анаеробне зброджування вуглеводів, як відомо, супроводжується декарбоксилюванням піровиноградної кислоти. Зброджування тих амінокислот, перетворення яких йде через стадію декарбоксилювання піровиноградної кислоти, також супроводжується утворенням СО2. Процес декарбоксилювання відбувається і в багатьох інших випадках, не пов'язаних з утворенням піровиноградної кислоти. По іншому стоїть питання у тому випадку, коли відомі схеми розкладу речовин не передбачають утворення СО2. При розкладі вищих жирних кислот, наприклад, це не повинно відбуватись, якщо врахувати, що основний шлях їх розкладу — β-окиснення. Між тим метан утворюється при зброджуванні будь-якого субстрату і більше всього при розкладі саме вищих жирних кислот. Як же у цьому випадку пояснити хімізм метаногенезу? Цілком очевидно, що у всіх випадках, незалежно від того, яка речовина є субстратом, що споживається, повинно бути джерело СО2. Складність відповіді на дане питання полягає, вірогідно, у тому, що механізм анаеробного розкладу речовин до цих пір остаточно не вивчений, у той час як аеробні процеси вивчені у всіх подробицях. Прийнято вважати, що анаеробний розклад речовин здійснюється тими ж шляхами, але враховуючи відсутність кисню воно йде не до кінця. До якої саме стадії — ми добре знаємо у випадку розкладу вуглеводів. В організмі людини і тварин анаеробний розклад вуглеводів закінчується утворенням молочної кислоти, у мікроорганізмів — утворенням різноманітних органічних кислот, спиртів, альдегідів, кетонів і т. д. Питання про анаеробне перетворення інших речовин до цих пір особливо не цікавило. Є підстави вважати, що хімізм перетворення речовин до циклу трикарбонових кислот у всіх випадках один і той самий. Цей хімізм добре відомий. Залишається встановити, на якій проміжній стадії перетворення речовин при метановому бродінні відбувається утворення СО2 і наступне відновлення його в метан. Такий проміжний продукт, безсумнівно, повинен бути загальним для хімізму розкладу всіх речовин. Одним з центральних метаболітів у всіх випадках, як відомо, є піровиноградна кислота. Однак розклад речовин не завжди супроводжується її утворенням. Більш загальним проміжним метаболітом є ацетил-КоА. Аеробний розклад вуглеводів, вищих жирних кислот і багатьох амінокислот проходить стадію ацетил-КоА, який піддається подальшому окисненню у циклі трикарбонових кислот. Ряд амінокислот вступає у цикл через піровиноградну кислоту, як наслідок, їх розклад проходить і стадію ацетил-КоА. Деякі амінокислоти залучаються в цикл через дикарбонові кислоти, у цьому випадку також є можливість перетворення їх в ацетил-КоА. Таким чином, можна сказати, що універсальним проміжним прод
|