Познание сущности кредита предполагает раскрытие его основы. В современной экономической литературе «основа» трактуется неодинаково. Часто под этим словом понимается та база, на которой зарождаются и развиваются кредитные отношения. Таким материальным основанием кредита в наиболее общем виде обычно считается товарное производство, более конкретно - кругооборот и оборот капиталов субъектов производства. Подобное толкование основы кредита как его базы, его материальной причины имеет определенный смысл, ведь с терминологической точки зрения это слово означает основание, фундамент. При такой постановке вопроса очевидно и другое - анализ как бы смещается в сторону выяснения причин, которые вызывают образование кредита, нас же интересует сущность кредита. Определенные плоскости анализа возникают и в том случае, если основа понимается как правила, регулирующие управление кредитным процессом. К ним довольно часто относят принципы кредитования (срочность кредита, его материальная обеспеченность), которые большинство авторов рассматривают как надстроечные элементы, как способы, с помощью которых обеспечивается реализация сущности кредитных отношений. Подобное направление анализа имеет полное право на существование, так как в системе кредитования должно быть некое ядро, которое предопределяет все другие составляющие элементы системы. В этом смысле принципы кредитования можно назвать основой, а все другие составляющие его механизма - обоснованными. Однако для целей анализа сущности подобный подход оказывается недостаточным. При таком понимании основа (как правило, способы кредитования) является объективированной, но не объективной категорией. В данном случае интерес представляет не процесс организации кредитных отношений, а их базисная сторона. Основа кредита - это наиболее глубокая часть сущности кредитных отношений, это то, на чем «держится» сущность и чем она определяется. Можно также предположить, что она составляет главное в сущности, определяющее свойство кредита, влияющее на все другие свойства. Определяющим звеном, главным в сущности кредита, некоторые авторы иногда считают уплату ссудного процента. На первый взгляд для этого действительно есть серьезные причины: ссудный процент неизбежно существует рядом с кредитом, без него кредитная сделка представляется противоестественной. Кажется, что без процента нет и кредита. В одной из книг начала XX в., посвященных кредиту и банкам, приводятся примеры, как один крестьянин передавал телегу во временное пользование. Является ли, спрашивает автор книги, такая передача вещи во временное пользование кредитом? Ведь здесь имеет место все - и передача, и возвращение вещи через определенный период времени. Тем не менее кредитом, считает автор, здесь можно назвать лишь такую сделку, когда крестьянин возвращает не только телегу, но и в виде платы кладет туда мешок овощей. В современном хозяйстве ссудный процент как бы становится атрибутом кредита, становится его свойством. Вместе с тем ссудный процент порождается не самим кредитом, а круговращением стоимости, которое свойственно капиталу. Это свойство не кредита, а использования средств как капитала. Ссудный процент выражает другое экономическое отношение, отличное от кредита, служит другой экономической категорией, возникающей при определенных условиях. Кредит является категорией обмена, в то время как ссудный процент выступает распределительной категорией. В истории развития кредита существуют многочисленные примеры беспроцентных ссуд. В римском праве, например, кредиты подразделяются на ссуды, по которым уплачивался процент, и ссуды, по которым процент не взыскивался. В современном хозяйстве беспроцентными зачастую выступают дружеские, личные кредиты (кредиты друзьям, близким знакомым, родственникам). Даже современный банковский кредит может приобретать беспроцентный характер (кредиты инсайдерам -акционерам банка и др.). Беспроцентными в особых случаях могут быть и международные кредиты, предоставляемые в порядке помощи развивающимся странам. Означает ли это, что современный кредит может существовать без ссудного процента? Видимо, нет. Признаком современной системы кредитования служит взыскание процента по ссудам. Связано это в том числе и с тем, что банковский кредит предоставляется главным образом на базе привлеченных средств, которые для банка выступают как платные ресурсы. Банки как коммерческие предприятия экономически не могут покупать ресурсы за плату, а продавать свои кредиты на бесплатной основе. Платный характер кредита порождает его движение как капитала. Поэтому и взыскание ссудного процента становится неотъемлемым правилом современной системы кредитования. Исходя из представленных суждений, нетрудно заметить, что основа кредита превращается в наиболее принципиальный вопрос, от решения которого зависит понимание сущности кредита. На наш взгляд, более правильно считать, что основой кредита является возвратность. Возвратность кредита, вне зависимости от стадий движения ссужаемой стоимости, выступает всеобщим свойством кредита. В процессе анализа кругооборота ссужаемой стоимости видно, что возвратность пронизывает все движение кредита, все стадии этого движения, будь то размещение и получение кредита или использование кредита, высвобождение стоимости, либо передача стоимости от заемщика к кредитору. Возвратность представляет собой специфическое свойство, определяющую черту, присущую кредитным отношениям. Возвратность не возникает сама по себе: она базируется на материальных процессах, на завершении кругооборота стоимости. Однако завершение кругооборота стоимости - это не возврат, высвобождение стоимости лишь подготавливает почву для возврата. Возврат кредита (в отличие от финансов) наступает в тот момент, когда высвободившиеся средства дают возможность ссудополучателю вернуть денежные средства, полученные во временное пользование. Возвратность выражает двусторонний процесс, она одинаково важна как для кредитора, так и для заемщика. Для кредитора ссужение стоимости - это не акт дарения; кредитор только потому и дает деньги (ценность) взаймы, что предполагает неизбежность их обратного притока. Для заемщика получение кредита - это также не дар, ему необходимо так использовать ссуду, чтобы обеспечить своевременное высвобождение стоимости и ее возврат. При возвратном движении стоимости важна и юридическая сторона. Собственность на стоимость, передаваемую на определенный срок, не переходит от кредитора к заемщику. Собственником в кредите остается первоначальный кредитор, заемщик становится лишь временным владельцем ссужаемых средств. Возвратность кредита - как бы возвратность в квадрате. Первоначально ссуженная стоимость возвращается, совершая круговращение в хозяйстве заемщика, высвободившиеся в результате такого круговращения средства заемщик передает затем своему кредитору. Подобное движение ссуженной стоимости существенно отличает кредит от других экономических категорий. Оно имеет принципиальное значение для практики: банки, аккумулирующие временно свободные ресурсы, не могут распоряжаться ими как своими капиталами. Собственником денежных средств, которые размещает банк на кредитной основе, остаются другие юридические и физические лица. Поэтому каждое решение о выдаче кредита должно соотноситься с тем, может ли банк получить обратно денежные средства, которые он разместил в виде кредита. Возвратность - объективное свойство. Это означает, что общество не может ее отменить, не изменив суть сделки. Лишь на базе объективных качеств кредитной сделки можно эффективно управлять кредитными потоками. Банк не имеет права размещать аккумулированные ресурсы на безвозвратной основе, так как он должен возвратить их истинным владельцам. Возвратность, свойственная кредиту, получает юридическое закрепление в соответствующем договоре, который заключают участники кредитной сделки и который фиксирует соглашение сторон, юридически закрепляет необходимость возвращения временно позаимствованной стоимости. На этом анализ основы кредита можно было и завершить, если бы не одна немаловажная проблема. Дело в том, что у кредита есть и социально-экономическая сторона. Как отмечалось ранее, кредит является порождением общественных потребностей. В этом смысле социально-экономическая основа кредита заложена в его общественном характере. Вне зависимости от типа общественно-экономических формаций кредит служит их целям. Кредит способствует реализации только таких общественных целей, при которых: • обеспечивается возвращение ссужаемой стоимости; • интересы кредитора и заемщика совпадают; • не допускается превалирование интересов одной из сторон кредитных отношений. Экономическая и социально-экономическая стороны основы кредита тесно взаимосвязаны друг с другом. Из истории кредита можно привести многочисленные примеры, когда акценты на потребности заемщика в ущерб интересам кредитора в конечном итоге приводили к задержке и даже потере возврата ссуд. Стремление кредитора заработать на заемщике, поглотить не только прибавочный, но и необходимый продукт (например, при ростовщическом кредите) приводило к разрушению экономики, фактическому свертыванию данной формы кредитных отношений. В результате анализа основы кредита его сущность можно определить как движение стоимости на началах возвратности в интересах реализации общественных потребностей. Данная характеристика кредита не исключает других определений его сущности. Сущность как выражение специфического имеет немало разнообразных оттенков. Уместить их в одном определении представляется затруднительным. В связи с этим определений сущности кредита может быть несколько, и каждое из них может выражать то или иное его качество, ту или иную деталь, характерную для его структуры, состава участников, стадий движения и основы кредита. Так, можно предположить, что кредит - это: • взаимоотношения между кредитором и заемщиком; • возвратное движение стоимости; • движение платежных средств на началах возвратности; • движение ссуженной стоимости; • движение ссудного капитала; • размещение и использование ресурсов на началах возвратности; • предоставление настоящих денег взамен будущих денег и др. Преимущество этих и других определений состоит в том, что каждое из них охватывает ту или иную сторону сущности кредита. Недостаток их заключается в том, что каждое из них в известной степени является ограниченным, не выражает всей сущности, не сможет поэтому претендовать на исчерпывающую характеристику. Было бы спорным при этом, если бы акцент делался на одном из них, в ущерб другим глубинным качествам кредитных отношений. Неверными выглядят и попытки выразить суть кредитных отношений через вскрытие их внешних качеств. В связи с этим вызывают возражения такие трактовки кредита, как: 1) денежные отношения; 2) формы распоряжения средствами; 3) аккумуляция и перераспределение временно свободных денежных средств. Определение кредитных отношений как денежных отношений исключают другие, неденежные формы кредита. Кредит как экономическая категория выражает свою суть независимо от того, в какой форме он выступает. Предоставление кредита в товарной форме не подтверждает его характеристику только как денежных отношений. В трактовке кредита как формы распоряжения средствами исчезает специфический элемент, затрагивающий сущностные черты этой экономической категории. Распоряжение ресурсами свойственно не только кредитным отношениям - распоряжаться можно и собственными средствами предприятий и организаций, а также финансами; данное качество, следовательно, не только не выделяет кредит из ряда других экономических отношений, но, напротив, как бы «растворяет» его в их массе. Наконец, нет достаточных оснований и в определении кредита как формы аккумуляции и перераспределения временно свободных денежных средств. Аккумуляция не выражает сущностный атрибут движения ссуженной стоимости. Накопление ресурсов, их аккумуляция в целях предстоящих затрат может осуществляться не столько в интересах их предоставления на началах кредита, сколько для использования накопленных ресурсов на нужды собственного производства, где они выступают не как полученные в кредит, а как собственные средства. Факт аккумуляции ресурсов не входит органически в понятие кредита, не может характеризовать сущность кредитных отношений. Перераспределение представляет собой не суть экономических отношений, а лишь одно из проявлений их сути. Не претендуя на полное выражение сущности кредита, можно было его охарактеризовать как передачу кредитором ссуженной стоимости заемщику для использования на началах возвратности и в интересах общественных потребностей
Вопросы для самоконтроля
Каковы факторы, обусловливающие необходимость кредита? 2. Какую роль в кредите играет доверие? 3. Чем кредит отличается от найма рабочей силы? 4. Почему страхование не является кредитом? 5. Чем кредит отличается от финансов? 6. Что важно проанализировать при раскрытии сущности кредита? 7. Что такое структура кредита? 8. Каковы стадии движения кредита? 9. Что такое основа кредита и в чем она выражается? 10. Каковы определения сущности кредита? 11. Какие определения сущности кредита дискуссионные?
Эволюция нервной системы хордовых
Нервная система хордовых во многом отличается от строения нервной системы у беспозвоночных животных. Хотя у высших хордовых (позвоночных) центральная нервная система подверглась значительной дифференциации, она, в сущности, сохранила организацию по принципу нервных стволов. Наряду с этим у позвоночных имеется автономная симаптическая нервная система, организованная по ганглионарному принципу.
Уникальность нервной системы хордовых особенно ярко проявляется в её онтогенезе.
У всех хордовых на стадии гастулы зачаток нервной системы располагается на спинной стороне зародыша, образуя нервную пластинку над зачатком хорды. В процессе нейруляции боковые края нервной пластинки начинают утолщаться и приподниматься в виде пары нервных (или медуллярных) валиков, так что средняя часть нервной пластинки оказывается вогнутой, - так формируется нервный желобок, тянущийся вдоль спинной стороны зародыша. Дальнейшее разрастание нервных валиков кверху и к средней линии завершается их смыканием друг с другом – желобок превращается в нервную трубку. Клетки покровной эктодермы, примыкающие сбоку к нервному желобку, интенсивно размножаются (пролиферируют) в направлении к средней линии, и эктодерма смыкается над нервной трубкой. На границе между медуллярными валиками и покровной эктодермой обособляются группы клеток, которые после смыкания нервной трубки располагаются над ней (и под эктодермой) в виде продольной полоски, называемой ганглионарной пластинкой (у некоторых позвоночных она имеет вид сжатого с боков нервного гребня).
Из нервной трубки развивается центральная нервная система (головной и спинной мозг у позвоночных). Ганглионарная пластинка распадается на отдельные клетки (мезенхиму), которые мигрируют в различных направлениях и образуют ганглии спинно - и черепномозговых нервов, ганглии симпатической автономной системы, а также пигментные клетки и целый ряд скелетных и мышечных структур головы.
Такое развитие нервной системы хордовых в онтогенезе хордовых, наличие в ней полости (невроцеля) и её расположение на спинной стороне тела над кишечником и осевым скелетом резко контрастирует с соответствующими характеристиками нервной системы высших групп беспозвоночных, у которых нервная система располагается на брюшной стороне тела под кишечником.
Эмбриональное развитие нервной системы хордовых из спинной нервной пластинки, а также ещё одна своеобразная её черта – светочувствительность (проявляющаяся у позвоночных в развитии глазных пузырей из выпячиваний передней части нервной трубки) позволили немецкому эмбриологу Т. Бовери обосновать гипотезу, предполагающую эволюционное возникновение нервной системы хордовых из особого нервно-рецепторного комплекса – так называемого аборального органа, имеющегося у гребневиков, а также у личинок многих групп высокоразвитых многоклеточных животных, в том числе иглокожих и полухордовых, возникших от общего корня с хордовыми. Аборальный орган расположен на верхнем полюсе тела и представляет собой комплекс, состоящий из нервного узла, стациоста (орган равновесия), примитивного глазка и султана чувствительных ресничек. Причины замены у предков хордовых первоначальной нервной системы (гомологичной нервным системам большинства групп беспозвоночных) вторичной, развивавшейся из аборального нервного комплекса, пока неясны. Может быть, они связаны с особенностями ориентации тела у предков хордовых при переходе их от плавания в толще воды к ползанию по поверхности морского дна.
У бесчерепных (низшие хордовые) морфогенез нервной системы, в сущности, завершается описанным выше состоянием: центральная нервная система у взрослых ланцетников представлена нервной трубкой, на спинной стороне которой даже сохраняется продольная перемычка между соприкоснувшимися, но не слившимися друг с другом медуллярными валиками (строго говоря, это даже не трубка, а нервный желобок с сомкнутыми краями). У личинок ланцетников передний конец нервной трубки связан с внешней средой отверстием (невропором), на месте которого позднее формируется обонятельная ямка. Внутренняя поверхность нервной трубки выстлана у ланцетника слоем клеток, сохраняющих эпителиальное строение. Среди них рассеяны более крупные нервные клетки и так называемые глазки Гессе – примитивные светочувствительные органы, с помощью которых ланцетник может определить направление падающего света и в какой-то мере оценить изменения его интенсивности (что важно для этих животных, ведущих «роющий» образ жизни в осадках на морском дне). Свет достигает глазков Гессе через полупрозрачные ткани животного. Кажущееся странным расположение светочувствительных органов внутри центральной нервной системы хорошо согласуется с предполагаемым происхождением последней от нервно – рецепторного комплекса, располагавшегося у предков хордовых на поверхности тела: при сворачивании нервной пластинки в трубку её поверхность, первоначально обращенная кнаружи, оказывается обращенной внутрь, к невроцелю.
В самом переднем конце нервной трубки ланцетника невроцель несколько расширен. Дно этой области образует так называемый инфундибулярный орган, представляющий собой небольшой выступ, в котором невроцель выстлан высокими цилиндрическими сенсорными клетками, несущие чувствительные волосковидные придатки. От нервной трубки в соответствии с сегментами туловищной мускулатуры (миомерами) поочередно отходят спинные и брюшные нервы, которые у ланцетников не объединяются в общий спинномозговой нерв, оставаясь самостоятельными. Спинные нервы образованы отростками нервных клеток, связанных с рецепторами в покровах тела и во внутренних органах, а также аксонами висцеральных двигательных нейронов. Эти нервы выходят из нервной трубки на границе между двумя миомерами и идут к покровам по перегородке между ними, затем висцеральная ветвь возвращается от покровов к внутренним органам. Спинномозговых ганглиев у ланцетника нет. Брюшные нервы представлены пучками нервных волокон (не образующими единого ствола), выходящими против миомеров и сразу в них проникающими.
В целом среди современных хордовых бесчерепные обладают наиболее примитивной нервной системой. Вероятно, близкое к этому уровню состояние было исходным для развития типов нервной системы, характерных для двух других подтипов хордовых – оболочников и позвоночных
У асцидий – наиболее характерных представителей подтипа оболочников – на соответствующих стадиях онтогенеза также развивается нервная трубка, расположенная в длинном хвостовом отделе личинок над хордой и кишечником. Передний конец нервной трубки несколько увеличен и образует так называемый мозговой пузырёк, с которым связаны рецепторы – многоклеточный светочувствительный глазок и одноклеточный статоцист. На более поздних стадиях онтогенеза асцидий происходит регрессивный метаморфоз (связанный с переходом взрослых животных к «сидячему» образу жизни на морском дне), в процессе которого основная часть нервной трубки атрофируется, и у взрослого животного сохраняется лишь надглоточный, или мозговой, ганглий, возникающий из спинной стенки мозгового пузырька личинки и расположенным над входом в глотку (между ротовым и клоакальным сифонами). Из брюшной стенки личиночного мозгового пузырька у взрослых асцидий развивается подмозговая железа, представляющая собой небольшой мешковидный орган, связанный с мерцательной ямкой (хеморецептором) в крыше глотки. Подмозговую железу считают гомологом инфунибулярного органа ланцетника и нейрогипофиза позвоночных.
Нервная система представителей двух других классов оболочников – сальп и аппендикулярий – в принципе сходна с таковой у асцидий (у сальп – со взрослыми асцидиями, у аппендикулярий – с личинками асцидий).
Дальнейшее прогрессивное развитие нервной системы произошло в эволюции высших хордовых – подтипа позвоночных, что, несомненно, было связано с общей активизацией образа жизни этих животных, древнейшие представители которых освоили активное плавание в толще воды на различных глубинах.
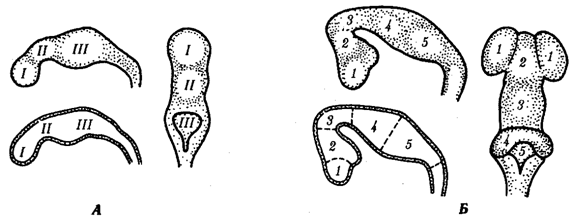
В онтогенезе позвоночных неравномерный рост разных частей головного отдела нервной трубки приводит к развитию нескольких вздутий (пузырей), разделенных перехватами. Первоначально развиваются вздутия (невромеры), примерно соответствующие зародышевым сегментам мезодермы (сомитам). Однако вскоре на их месте формируются более крупные образования – три первичных мозговых пузыря (см. рис.1.2): первичный передний, средний и ромбический мозг (им соответствуют I-III, IV-V и VI-XI невромеры). Предполагают, что эта стадия онтогенеза соответствует первичной дифференциации головного мозга древних позвоночных на центры обонятельной, зрительной и статоакустической функций. Неравномерный рост разных участков формирующегося головного мозга продолжается далее и приводит, во-первых, к разделению первичного переднего мозгового пузыря на два – собственно передний, или конечный, мозг и промежуточный мозг, во-вторых, к дифференциации ромбического мозгового пузыря на задний мозг (развивающийся как вырост нервной стенки первого) и продолговатый мозг. Это стадия пяти мозговых пузырей (см. рис.).
Продолговатый мозг сзади продолжается в спинной мозг, который сохраняет внешне малодифференцированное строение нервной трубки. Однако внутреннее строение спинного мозга (как, разумеется, и головного) у позвоночных подвергается сложной дифференцировке.Тела нервных клеток группируются вокруг невроцеля, образуя в совокупности серое вещество спинного мозга, у высших позвоночных напоминающее на поперечном срезе фигуру бабочки. «Крылья бабочки» образуют так называемые спинные и брюшные рога серого вещества. В спинных рогах располагаются нейроны, которые получают информацию от рецепторов, воспринимающих раздражения из внешней среды (соматическая чувствительная или соматосенсорная, функция). Ближе к основанию спинных рогов находятся висцеральные нейроны, получающие информацию от рецепторных клеток, которые находятся во внутренних органах. Брюшные рога образованы телами соматических двигательных нейронов, контролирующих работу поперечнополосатой мускулатуры тела и конечностей. Наконец, в средней части «бабочки» серого вещества (у млекопитающих здесь образуются набольшие боковые рога) расположены висцеральные двигательные нейроны, под контролем которых находится мускулатура внутренних органов (преимущественно гладкие мышечные волокна в стенках органов пищеварения, дыхания, выделения). Вокруг «бабочки» серого вещества располагается белое вещество, образованное аксонами нервных клеток. Белый цвет этой области обусловлен миелинизированными оболочками аксонов. Здесь проходят проводящие пути, по которым информация передаётся внутри центральной нервной системы. Между спинными рогами серого вещества располагаются так называемые спинные столбы серого вещества – проводящие восходящие пути, по которому информация передаётся к головному мозгу. Соответственно, между брюшными рогами находятся брюшные столбы, образованные аксонами двигательных нейронов, нисходящими от головного мозга к мышцам тела. Наконец, по бокам спинного мозга располагаются боковые столбы, содержащие аксоны как восходящих, так и нисходящих путей. Спинные и брюшные столбы левой и правой стороны по средней линии в той или иной степени разделены узкими продольными спинной и брюшной бороздами, заполненными соединительной тканью.
В головном мозге картина дифференциации серого и белого вещества гораздо более сложна и менее правильна, чем в спинном. Серое вещество образует здесь различные ядра, окруженные проводящими путями (белое вещество), а также в некоторых отделах кору (слой серого вещества у поверхности мозга).
В онтогенезе после стадии пяти мозговых пузырей продолжается дальнейшая дифференцировка головного мозга.
Мозговые пузыри в эмбриогенезе мозга человека. А — стадия трех мозговых пузырей; Б — стадия пяти мозговых пузырей
Передний мозг, сохраняющий у всех позвоночных функцию нервного центра обонятельного анализатора, впереди разделяется продольно на пару обонятельных долей. Позади них дифференципуются полушария, где у высших позвоночных локализуются высшие ассоциативные центры. Основная масса нервной ткани переднего мозга у низших позвоночных локализована в нижней части полушарий, где находятся основные ганглии, или полосатые тела. Верхняя часть стенок полушарий, первоначально не содержащая нервной ткани, называется первичным плащом (архипаллиум). Невроцель в области переднего мозга разделен на пару (левый и правый) желудочков и полушарий и обонятельных долей; в задних отделах головного мозга невроцель остаётся непарным.
В морфогенезе промежуточного мозга его стенки и дно образуют 4-5 пузыревидных выпячиваний, растущих в разных направлениях. По бокам развиваются глазные пузыри; позднее их боковые стенки внутрь и плотно прилегают ко внутренним – глазные пузыри преобразуются в глазные бокалы, двуслойные стенки которых образуют сетчатку и пигментную оболочку глаз. Два аналогичных пузыревидных светочувствительных выпячивания растут от промежуточного мозга кверху, располагаясь друг за другом в срединной плоскости. Это теменной и пинеальный органы. Они не являются глазами в обычном смысле слова. Восприятие ими света используется для регуляции функций различных органов в соответствии с фотопериодом в суточном и годовом циклах. Активную роль в этой регуляции играет гипофиз – одна из важнейших эндокринных желез позвоночных, расположенная у дна промежуточного мозга. Её задняя доля – нейрогипофиз – образуется из непарного выроста дна промежуточного мозга, называемого воронкой. В нижней стенке промежуточного мозга расположен гипоталамус, включающий так называемый серый бугор впереди воронки, нижние доли – в её боковых стенках и сосудистый мешок – позади. Гипоталамус образован нервными и нейросекреторными клетками, а также проводящими путями, связывающими передние и задние отделы головного мозга и верхние и нижние придатки промежуточного мозга. Здесь находится своего рода перекресток центральных проводящих путей головного мозга и нейроэндокринный центр, регулирующий размножение и важнейшие вегетативные функции организма (обмен веществ, температура тела, кровяное давление, дыхание, пищеварение, поддержание постоянства внутренней среды организма, т.е. гомеостаз). Гипоталамус вместе с гипофизом образуют единый нейроэндокринный комплекс, называемый гипоталамо-гипофизной системой. Невроцель в области промежуточного мозга образует третий желудочек, верхняя стенка которого представляет собой перепонку, не содержащую нервных клеток.
Крыша среднего мозга вздута в виде пары зрительных долей (в глубине которых располагаются не только зрительные, но и слуховые ядра). Невроцель в этой области у низших позвоночных довольно велик, а тетрапод представлен лишь узким каналом (так называемый сильвиев водопровод), связывающим широкие и открытые сверху третий и четвертый желудочки. Последний, называемый также ромбовидной ямкой, представляет невроцель продолговатого мозга.
Задний мозг у большинства позвоночных образован мозжечком, который развивается в онтогенезе как верхняя поперечная складка впереди четвёртого желудочка. Мозжечок участвует в координации движений и регуляции мышечного тонуса. У млекопитающих к заднему мозгу присоединяется также варолиев мост – часть ствола мозга между продолговатым и средним мозгом. Здесь располагаются проводящие пути и ряд ядер серого вещества, в том числе ядра V-VIII пар черепномозговых нервов и некоторые структуры так называемой ретикулярной формации, продолжающейся в среднем и продолговатом мозгу. Структуры этой формации являются системой двигательного контроля. В продолговатом мозгу расположены также нервные центры, контролирующие дыхание, кровообращение и обмен веществ.
Вообще необходимо подчеркнуть, что границы между отделами головного мозга в значительной степени условны: они плавно переходят один в другой, и мозг анатомически представляет собой единый орган. Кроме того, и функции разных отделов центральной нервной системы сложно переплетены. Большинство функций обеспечиваются взаимодействием нейронов, локализованных в нескольких ядрах различных отделов мозга. Поэтому функциональная морфология нервной системы крайне сложна; её изучение требует анализа многообразных взаимосвязей разных нервных центров, формирующих сложноорганизованные функциональные системы, которые перекрываются друг с другом по всему представительству в разных отделах мозга. Современные представления в этом отношении значительно отличаются от таковых науки прошлого, когда предполагали, что в головном мозге можно обнаружить своего рода «мозаику» из нервных центров, каждый из которых отвечает за определённую соматическую или висцеральную функцию.
Для периферической нервной системы позвоночных характерен то же общий план строения, который наблюдается у бесчерепных. Отличия заключаются в появлении спинномозговых ганглиев, в которых расположены тела первичных чувствительных нейронов (как соматических, так и висцеральных), и в объединении соматических двигательных волокон каждого сегмента в пару брюшных корешков.
Подтип: Бесчепные (Acrania) Ланцетник: Центральная нервная система не дифференцирована на головной и спинной мозг. ВЧ связи с отсутствием головного мозга нет черепа. Органы чувств развиты слабо: имеются разбросанные по поверхности тела осязательные клетки, светочувствительные образования глазки Гессе, расположены в стенках нервной трубки, и лежащая в головном отделе на поверхности тела обонятельная ямка. Подтип: Черепные (Craniata) Класс МиногиМинога: ЦНС делится на спинной и головной мозг (он весьма мал). Головной мозг имеет отделы характерные для всех позвоночных животных:Передний мозг, промежуточный мозг, средний мозг, мозжечок и продолговатый мозг. Примитивность – все отделы лежат в одной плоскости и не налегают друг на друга. Передний мозг относительно других отделов очень невелик. Хорошо заметны обонятельные доли – выпячивание его передней части. Полость внутри него двураздельная, что свидетельствует о формировании боковых желудочков. Крыша переднего мозга эпителиальная.Промежуточный мозг хорошо виден сверху. Сверху его находятся два органа – теменной (имеет пигментные и светочувствительные клетки) и эпифизальный. На нижней стороне промежуточного мозга находится гипофиз.Средний мозг развит слабо. Небольшие зрительные доли развиваются в его боковых стенка, между ними из-за недоразвития крыши имеется отверстие. Мозжечок очень невелик и имеет вид валика, который ограничивает полость продолговатого мозга – ромбовидную ямку спереди и сверху. Недоразвитость мозжечка связана с простотой движения. Продолговатый мозг имеет относительно крупные размеры и посепенно переходит в спинной мозг. Полость его спереди сообщается с полостью среднего мозга, а сзади постепенно переходит в спинно – мозговой канал. От головного мозга миноги отходят 10 пар головных нервов В связи со слабым развитием затылочного отдела черепа нервы 9 (языкоглоточный) и 10 пары (блуждающий) отходят от мозга вне черепа. Спиной мозг лежит над хордой в спинно-мозговом канале и имеет плоскую лентовидную уплощённую форму. Надкласс рыбы (Pisces) Класс: Хрящевые рыбы (Chondrichthyes)Акула: В отличие от миноги передний мозг и мозжечок имеют крупные размеры. Обонятельные доли переднего мозга относительно велики. Нервная ткань переднего мозга выстилает не только его бока, дно, но и крышу. Поверхность мозжечка образует систему извилин.На крыше промежеточного мозга небольшое утолщение – эпифиз, снизу гипофиз – железы внутренней секреции. Средний мозг так же хорошо развит.Происходит наложение мозжечка на значительную часть зрительных долей среднего мозга спереди, а сзади мозжечок налегает на часть продолговатого мозга. Класс: костные рыбы (Osteichthyes)Хорошо развиты обонятельные нервы.Промежеточный мозг сверху прикрыт большими полушариями и средним мозгом. Средний мозг у костных раб заметно крупнее остальных отделов. Сверху заметны две крупные зрительные доли.Мозжечок также достаточно велик, особенно у подвижных видов рыб. Он значительно прикрывает продолговатый мозг. Продолговатый мозг удлинён и постепенно переходит в спинной мозг. Сверху заметна ромбовидная ямка – четвёртый желудок мозга. При рассмотрении головного мозга снизу явственно видны передний мозг с обонятельными долями, округлый вырост на промежуточном мозге – нипофиз и перед ним перекрёст зрительных нервов (хиазма) Спинной мозг лежит в канале, образованном верхними дугами позвоков. Класс: Земноводные (Amphibia)Лягушка: Обладает рядом прогрессивных черт. Главным образом это касается переднего мозга, который у амфибий относительно крупнее, чем у рыб, полушария его полностью разделены, и нервное вещество выстилает кроме дна боковых желудочков также бака и крышу, т.е у амфибий имеется настоящий настоящий мозговой свод – архипалииум. Среди костных рыб настоящий мозговой свод свойствен лишь двоякодышащим. Впереди расположен передний мозг, состоящий из двух разделённых глубокой щелью удлиненных полушарий. Спереди от полушарий отходит большая обонятельная доля, от которой берут начало два обонятельных нерва. Позади переднего мозга находится промежуточный мозг. На его крыше располагается эпифиз (железа внутренней секреции). Средний мозг представлен в виде двух округлых зрительных долей. Сзади зрительных долей лежит слаборазвитый мозжечок (в связи с однообразностью движений). Сразу же за ним расположен продолговатый мозг с ромбовидной ямкой. Продолговатый мозг постепенно переходит в спинной мозг. Как и у большинства хордовых животных, нервная система пресмыкающихся представлена головным (из 5 отделов) и спинным мозгом. Головной мозг расположен внутри черепа. Ряд важных особенностей отличает головной мозг пресмыкающихся от головного мозга земноводных. Нередко говорят о так называемом зауропсидном типе головного мозга, присущем также и птицам, в отличие от ихтиопсидного типа у рыб и земноводных. Выделяют пять отделов головного мозга пресмыкающихся. Передний мозг состоит из двух больших полушарий, от которых отходят обонятельные доли. Поверхность больших полушарий абсолютно гладкая. В мозговом своде полушарий различают первичный свод — архипаллиум, занимающий большую часть
ТЕОРЕТИЧЕСКАЯ МЕХАНИКА Статика является частью теоретической механики, изучающей условия, при которых тело находится под действием заданной системы сил...
Теория усилителей. Схема Основная масса современных аналоговых и аналого-цифровых электронных устройств выполняется на специализированных микросхемах...
Понятие массовых мероприятий, их виды Под массовыми мероприятиями следует понимать совокупность действий или явлений социальной жизни с участием большого количества граждан...
ОЧАГОВЫЕ ТЕНИ В ЛЕГКОМ Очаговыми легочными инфильтратами проявляют себя различные по этиологии заболевания, в основе которых лежит бронхо-нодулярный процесс, который при рентгенологическом исследовании дает очагового характера тень, размерами не более 1 см в диаметре...

 Нервная система представителей двух других классов оболочников – сальп и аппендикулярий – в принципе сходна с таковой у асцидий (у сальп – со взрослыми асцидиями, у аппендикулярий – с личинками асцидий).
Нервная система представителей двух других классов оболочников – сальп и аппендикулярий – в принципе сходна с таковой у асцидий (у сальп – со взрослыми асцидиями, у аппендикулярий – с личинками асцидий).