Слуховая сенсорная система позвоночных животных
У всех позвоночных животных рецепция звуковых колебаний осуществляется специальными структурами, расположенными во внутреннем ухе, или лабиринте. Возникновение лабиринта как замкнутой системы, расположенной глубоко под кожей головы, рассматривают как значительный прогресс в эволюции органов слуха. У самых примитивных водных позвоночных — круглоротых — уже имеется хорошо развитый лабиринт. У первичноводных позвоночных, так же как и у некоторых животных, ведущих смешанный образ жизни (земноводных), кроме лабиринта в восприятии звуковых колебаний участвуют еще и органы боковой линии (см. разд. 4.4). Рецепторные структуры органов боковой линии и внутреннего уха осуществляют различные функции при обнаружении и опознавании акустических сигналов. Воспринимающие звуковые волны волосковые клетки лабиринта сгруппированы в виде макул, прикрытых желатиновидной отолитовой мембраной с одним крупным отолитом или многими мелкими отокониями и папиллами (от лат. papilla — сосочек), с односторонне прикрепленными кроющими образованиями — текториальными мембранами. В разных группах позвоночных слуховые функции выполняют различные макулы и папиллы. У рыб это саккулярная и лагенарная макулы. Амфибиальная и базилярная папиллы выполняют слуховые функции у амфибий, новая базилярная папилла — у рептилий, птиц и млекопитающих. При этом у последних базилярная папилла обособляется в виде улитки — прямой или спирально закрученной, образуя специализированный орган слуха. Рецепторные клетки, заключенные в улитку, функционируют как приемники давления и нуждаются в дополнительных системах, предварительно обрабатывающих сигналы и оптимизирующих их восприятие. Такие системы становятся совершенно необходимыми в связи с выходом позвоночных на сушу и существованием в воздушной среде, в которой их тела становятся непроницаемыми для звука. Этим объясняется появление в слуховом аппарате наземных позвоночных звукопоглощающих, звукопередающих, а позднее и звукоорганизующих структур, представленных в среднем и наружном ухе. Эволюция слуховой системы позвоночных. Возникновение у животных специфического приемника звука неразрывно связано с водной средой, в которой на протяжении многих геологических эпох развивалась жизнь, предшествующая появлению разнообразных биологических форм на суше. У рыб орган слуха представлен лишь внутренним ухом, которое размещается частично в боковых стенках черепа и большей частью на его внутренней поверхности. Уже в 30—х гг. XX века Г. Фриш доказал, что слуховую функцию несет нижняя часть лабиринта — саккулюс, тогда как утрикулюс и полукружные каналы участвуют в поддержании равновесия. Волны звукового давления ближнего поля, распространяющиеся в водной среде, легко передаются через ткани и жидкости в теле рыбы и, таким образом, создают колебательные движения макулы, покоящейся на твердом субстрате. В результате реснички волосковых клеток, концы которых тесно прикреплены к отолиту с помощью текториальных тяжей, начинают двигаться одни относительно других. Силы, действующие на реснички, заставляют их сгибаться, что приводит к стимуляции волосковых клеток. Что касается волн давления, определяющих изменения дальнего поля, то, несмотря на их высокую амплитуду в определенных условиях, они непосредственно не возбуждают лабиринт, так как находятся вне предела чувствительности его сенсорных структур. Для их восприятия необходимо иметь прибор, который бы усиливал эти волны и переводил их, таким образом, в зону чувствительности лабиринтных структур. В качестве такого усиливающего воспринимающего прибора рассматривают плавательный пузырь. Плавательный пузырь имеет определенный резонанс в пределах 100—1000 Гц. Руководствуясь резонансными свойствами плавательного пузыря, можно считать, что рыбы чувствительны к волнам дальнего поля в диапазоне колебаний 0,1—1,0 кГц. В процессе эволюции у наиболее высокоорганизованных водных позвоночных развивались специальные образования, усиливающие колебания дальнего поля при передаче их к структурам лабиринта. К подобным образованиям относятся прежде всего веберов аппарат и другие функционально гомологичные приспособления. Соединение плавательного пузыря с внутренним ухом осуществляется с помощью веберова аппарата — четырех пар подвижно сочлененных косточек, являющихся аналогом системы косточек среднего уха млекопитающих, тогда как плавательный пузырь считают функциональным аналогом барабанной перепонки. При колебаниях плавательного пузыря движения его стенок через систему косточек передаются к непарному синусу, а затем к поперечному каналу и саккулюсу. Перемещение эндолимфы к сенсорному эпителию макулы осуществляется непосредственно за счет движения жидкости в поперечном канале. В этом случае отолит качается по своей длинной оси и возбуждает волосковые клетки макулы. Важнейший этап в эволюции слуховой системы позвоночных был связан с переходом от водного к наземному образу жизни. Это потребовало развития у животных специализированных структур, согласующих сопротивление проводимых по воздуху звуков с сопротивлением жидкостей внутреннего уха. Впервые по сравнению с рыбами в пределах класса земноводных, или амфибий уже у древних стегоцефалов (палеозой) появляется барабанная полость, образованная из щели между челюстной и гиоидными дугами. Несомненно существовала и барабанная перепонка, которая хорошо развита у современных бесхвостых земноводных. За исключением борнеосской лягушки, бесхвостые земноводные не имеют наружного слухового прохода и их барабанная перепонка располагается открыто по бокам головы. Аппарат среднего уха у них примитивен и представлен слуховым столбиком, который одним концом через хрящ соединен с барабанной перепонкой, а другим упирается в овальное окно, контактирующее с перилимфатической цистерной внутреннего уха. Перепончатый лабиринт внутреннего уха амфибий находится во взвешенном состоянии внутри костной слуховой капсулы. В целом перепончатый лабиринт вместе с заключенными в нем амфибиальной и базилярной папиллами является эндолимфатической системой, а окружающее его пространство заполнено перилимфатической жидкостью. Предполагают, что базилярная папилла имеет преимущественное значение при восприятии высокочастотных звуковых колебаний в воздухе, а амфибиальная папилла — низкочастотных в воздухе и воде. Эффективный диапазон частот, вызывающий изменение частоты дыхания у лягушек, находится в пределах 30—15 000 Гц. Слуховой аппарат рептилий устроен значительно сложнее амфибий и имеет ряд прогрессивных черт. Так, у гекконов, сцинков, ящериц и крокодилов появляется наружный слуховой проход, обладающий особыми кольцевыми мышцами, которые могут перекрывать или сильно суживать наружное слуховое отверстие. Барабанная полость среднего уха заполнена воздухом и широко открывается в гортань. В барабанную полость открывается отверстие, аналогичное окну улитки млекопитающих. Такая система проводит и усиливает звуковые воздушные колебания. Своеобразные изменения наблюдаются и в строении внутреннего уха. Базилярная папилла рептилий представляет собой совершенно новую структуру, лишенную непосредственного сходства с базилярной папиллой амфибий. Она располагается в улитковом канале, являющемся выростом саккулюса, и примыкает к лагенарной макуле. Существенная отличительная черта базилярной папиллы рептилий — расположение волосковых клеток на структурно дифференцированной базилярной мембране (рис. 4.28), что создает возможность для гармонического анализа звуков уже на уровне рецептора.
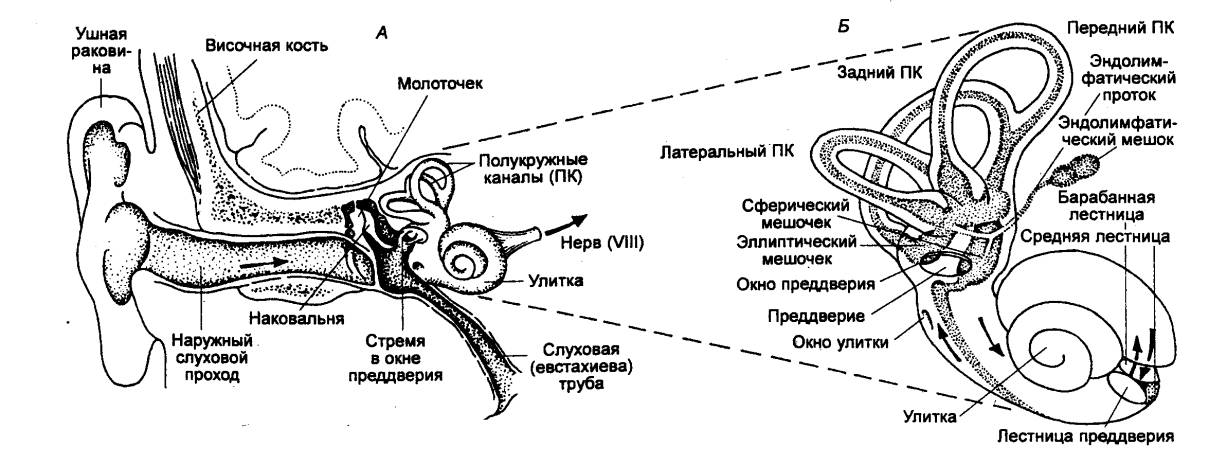
Поверхность папиллы покрывает текториальная мембрана. Благодаря такой организации стимуляция волосковых клеток происходит не за счет отклонения их ресничек (как у рыб и амфибий), а за счет движения тел самих клеток в результате изгибания базилярной мембраны под влиянием колебаний эндолимфы. Диапазон частот, воспринимаемых разными видами рептилий, составляет 50—10 000 Гц, причем максимальная чувствительность обнаруживается, как правило, в области частот ниже 1 кГц. У птиц наружный слуховой проход обычно короткий, большого объема, очень сложный по рельефу складок. Длительное время считали, что наружное ухо птиц является исключительно защитным образованием, однако показано, что оно несет совершенно определенную акустическую нагрузку и влияет на частотные характеристики слуха. Среднее ухо птиц представлено обширной тимпанальной полостью. Звукопередающий аппарат среднего уха состоит из хрящевой экстраколюмеллы (столбика) с тремя отростками на барабанной перепонке. На них фиксируется единственная мышца среднего уха — тимпанальная, которая берет начало от боковой затылочной кости вблизи мыщелка. Единственная слуховая косточка птиц представляет собой пластинку, оканчивающуюся диском, который упирается в овальное окно. Внутреннее ухо представлено короткой, слабо изогнутой трубкой. На базилярной мембране располагается вытянутый в длину спиральный орган, который состоит из опорных и рецепторных клеток, покрытых с поверхности текториальной мембраной. Дистальную часть улитки занимает лагена, тогда как в проксимальной части расположен собственно акустический рецептор — базилярная папилла. Диапазон частот, воспринимаемый птицами, довольно широк и простирается от 30 Гц до 30 кГц. Отмечается высокая степень частотного различения. Например, дифференциальный порог по частоте у волнистых попугайчиков в диапазоне 0,3—1,0 кГц составляет всего 0,3—0,7%, т. е. близок к возможностям человеческого слуха. Из всех позвоночных животных млекопитающие обладают наиболее развитым периферическим слуховым аппаратом. В каждом отделе наружного, среднего и внутреннего уха произошли существенные изменения по сравнению с рассмотренными классами, поэтому представляется целесообразным рассмотреть строение и функции периферических отделов слуховой системы млекопитающих более подробно. Слуховая система млекопитающих. Наружное ухо млекопитающих (рис. 4.29) включает ушную раковину и наружный слуховой проход. По своей форме наружное ухо несколько напоминает воронку с усложненной внутренней поверхностью ушной раковины за счет развития системы дополнительных клапанов и увеличения ее размеров. Воронкообразная форма наружного уха обеспечивает свойство направленности — улучшенное восприятие звуков, идущих с определенного направления, по сравнению со звуками, идущими с других направлений. Ушные раковины отсутствуют только у китообразных и многих ластоногих, что является специальным приспособлением к водному образу жизни.
Среднее ухо впервые представляет трехкосточковую звукопередающую систему, включая молоточек, наковальню и стремя, связанные с одной стороны с барабанной перепонкой, а с другой — с окном преддверия (овальным окном) внутреннего уха. Полость среднего уха, включая и водных животных, заполнена воздухом. Слуховые косточки выполняют двоякую роль. Их первая функция состоит в том, что они образуют систему рычагов, с помощью которых улучшается передача энергии колебаний из воздушной среды слухового прохода к перилимфе внутреннего уха. Благодаря тому, что площадь основания стремени, укрепленного в окне преддверия, значительно меньше площади барабанной перепонки, а также благодаря специальному способу сочленения косточек, действующих наподобие рычагов, давление на мембране овального окна оказывается примерно в 20 раз большим, чем на барабанной перепонке. Этот механизм увеличения давления является чрезвычайно целесообразным приспособлением, направленным на обеспечение эффективной передачи акустической энергии из воздушной среды в жидкую. Вторая функция заключается в способности системы косточек изменять характер движения при больших интенсивностях звука. Когда звуковое давление приближается к величинам порядка 120 дБ (над порогом слышимости), человек начинает ощущать покалывание в ушах. При таких интенсивностях стимула существенно меняется характер движения косточек, что резко снижает функцию среднего уха. В среднем ухе есть также специальный механизм, предохраняющий слуховой рецепторный аппарат от длительных звуковых перегрузок. Достигается это сокращением мышц среднего уха, которых у млекопитающих уже две: мышца, напрягающая барабанную перепонку, и стременная мышца. Рефлекторное сокращение этих мышц при действии звука большой интенсивности приводит к уменьшению амплитуды колебания барабанной перепонки, косточек среднего уха и, соответственно, к уменьшению звукового давления, передаваемого улитке. Внутреннее ухо (улитка) млекопитающего представляет собой спирально закрученный костный канал, приподнятый в области вершины. У человека улитка имеет 2,5 витка, у кошки — 3, а у однопроходных — всего 0,25 витка. В улитке млекопитающих (за исключением однопроходных) отсутствует лагенарная макула, и весь рецептор представлен базилярной папиллой, видоизмененной в спиральный (кортиев) орган.
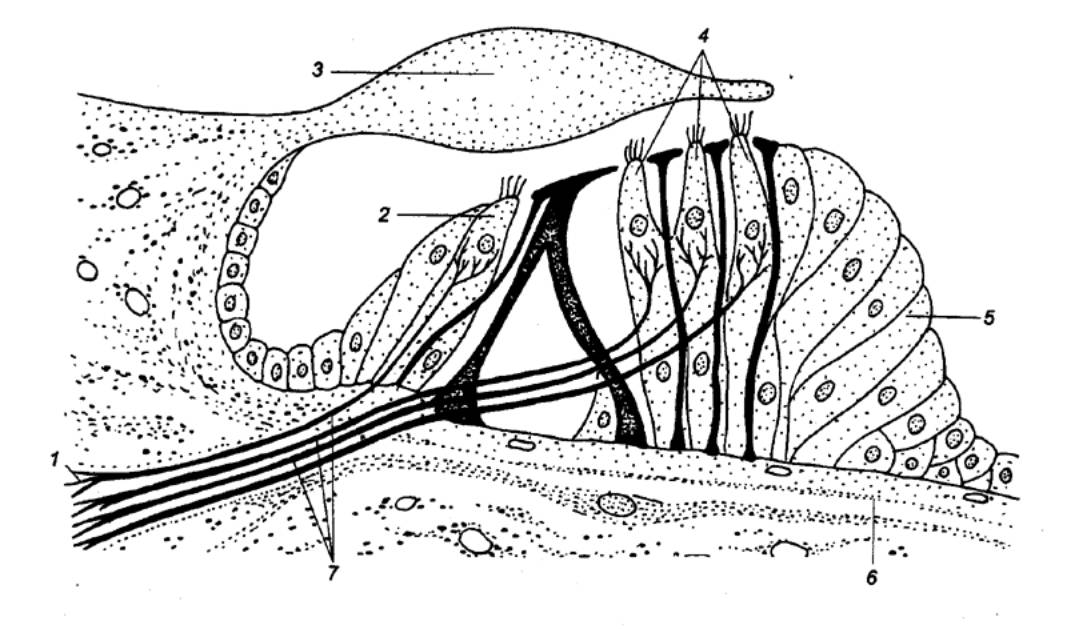
Внутри костной капсулы две мембраны (базилярная пластинка и преддверная мембрана, или мембрана Рейсснера) разделяют улитку на три лестницы: барабанную, улитковый проток (среднюю) и лестницу преддверия. Барабанная лестница и лестница преддверия заполнены перилимфой, улитковый проток — эндолимфой. На базилярной пластинке располагается спиральный орган (рис. 4.30), который включает два типа рецепторных клеток: один ряд внутренних и три—четыре ряда наружных волосковых клеток. Каждая рецепторная клетка увенчана пучком стереоцилий и в отличие от рептилий и птиц не имеет киноцилии (вместо нее располагается базальное тельце, или центриоль). Стереоцилий прикрепляются к нижней поверхности покровной мембраны. Волосковые клетки, так же как и у птиц, располагаются на опорных клетках, которые у млекопитающих подразделяются на клетки—столбы, клетки Хензена, наружные поддерживающие (Клаудиса) и наружные фаланговые (Дейтерса). Базилярная пластинка не одинакова по ширине: у человека вблизи окна преддверия ее ширина составляет 0,04 мм, а затем по направлению к куполу улитки, постепенно расширяясь, она достигает в конце 0,5 мм. Следовательно базилярная пластинка расширяется там, где улитка сужается. В базальной части спирального органа располагаются рецепторные клетки воспринимающие более высокие частоты, а в апикальной части (на вершине улитки) — клетки, воспринимающие только низкие частоты. Такой пространственный способ анализа частоты получил название принципа места. Однако представления о механизме, на котором основан такой способ кодирования, за последние сто лет претерпели существенные изменения. Еще в 1863 г. Г. Гельмгольц сформулировал резонансную теорию слуха, согласно которой разные частоты кодируются своим точным положением вдоль базилярной пластинки. Базилярная пластинка может действовать как набор поперечно натянутых эластичных резонирующих полос, подобных струнам рояля. Самые короткие из них в узкой части близ основания улитки резонируют в ответ на высокие частоты, а те, что лежат ближе к куполу, в расширенной части базилярной пластинки, — на самые низкие частоты. Эта теория основывалась на том, что базилярная пластинка натянута по ширине и механическая связь по ее длине отсутствует, т. е. колебание одной части мембраны не должно передаваться соседним участкам.
Однако эти исходные предпосылки были опровергнуты в 50—60—е гг. XX столетия Д. Бекеши. Им прежде всего было доказано, что базилярная пластинка не натянута в поперечном направлении и что она имеет механическую связь по всей длине. Поэтому он предложил свою новую теорию, которую обычно называют теорией бегущей волны. Бекеши установил, что базилярная мембрана жестче всего у основания улитки, т. е. там, где она уже. По направлению к куполу ее жесткость постепенно уменьшается. При колебаниях мембраны волны «бегут» от ее основания к куполу. Градиент жесткости мембраны всегда заставляет волны двигаться от окна преддверия и никогда в обратном направлении. Высокочастотные колебания продвигаются по базилярной пластинке лишь на короткое расстояние, а длинные низкочастотные волны распространяются довольно далеко. Первая, самая жесткая часть базилярной пластинки служит высокочастотным фильтром. Таким образом, энергия коротковолновых колебаний рассеивается, шунтируется, так что они затухают недалеко от основания, тогда как длинные волны проходят весь путь до вершины. Бегущая волна имеет наибольшую амплитуду на строго определенном участке мембраны в зависимости от частоты. И хотя сама волна движется, ее огибающая для данной частоты стационарна. Смещения пиков для высоких частот направлены к основанию, а для низких частот — к куполу улитки. Электрические явления в улитке, регистрируемые в ее средах при отсутствии звукового раздражения и возникающие при действии звукового стимула, можно подразделить на две группы. В первую входит постоянный эндолимфатический потенциал. Хотя он регистрируется при отсутствии звука, обнаружены его существенные изменения при колебаниях базилярной пластинки. Наличие эндолимфатического потенциала обеспечивает высокую поляризованность структур улитки, что приводит к высокой чувствительности механизма преобразования механической энергии в процесс возбуждения рецепторов. Вторую группу электрических явлений, возникающих в улитке при действии звука, составляют микрофонный и суммационный потенциалы. Микрофонные потенциалы впервые были обнаружены в спиральном органе кошки, а затем в органах слуха других млекопитающих. Они повторяют в ряде случаев форму звуковой волны. Амплитуда микрофонного потенциала наиболее велика в тех участках улитки, которым соответствует максимум колебаний базилярной пластинки при данной частоте тона. С помощью внутриклеточного отведения была зарегистрирована активность одиночной волосковой клетки. Она показала довольно острую настройку на характеристическую частоту. В отличие от микрофонного потенциала суммационный потенциал воспроизводит не форму звуковой волны, а ее огибающую. Различают положительный и отрицательный суммационные потенциалы. Считают, что отрицательный суммационный потенциал связан с внутренними, а положительный и микрофонный — с наружными волосковыми клетками. Заключительный этап деятельности улитки характеризуется возникновением импульсации в волокнах слухового нерва, иннервирующих рецепторные волосковые клетки. Характер этой иннервации достаточно сложен. Примерно 95% сенсорных слуховых волокон связаны только с внутренними волосковыми клетками, которые у млекопитающих составляют лишь около 20% всех волосковых клеток (у человека их всего 3500). Напротив, более многочисленные наружные волосковые клетки (у человека их более 20 000) связаны только с немногочисленными сенсорными волокнами. Таким образом, внутренние волосковые клетки обладают множественной иннервацией, которая, вероятно, обеспечивает большую надежность передачи в отличие от разветвляющейся иннервации многих наружных клеток от одного волокна, которая связывает активацию волокна одной волосковой клетки с одновременной активностью соседних. Полагают, что основной поток слуховых ответов идет через внутренние волосковые клетки, причем наружные клетки вносят определенный вклад в свойства сигналов. Кроме афферентных волокон, несущих сенсорную информацию, к волосковым клеткам подходят также и эфферентные волокна. Они идут от клеток ядра верхней оливы в стволе мозга и образуют синаптические связи с волосковыми клетками. Стимуляция этих волокон вызывает угнетение ответов волосковых клеток. Возможно, эти волокна каким—то образом защищают волосковые клетки от перераздражения, но в остальном их функции остаются неизвестными. Диапазон воспринимаемых частот в пределах класса млекопитающих весьма различен, но основной тенденцией является хорошо выраженная чувствительность к высоким частотам. Расширение полосы высокочастотных сигналов является результатом эволюции цепи слуховых косточек в среднем ухе млекопитающих по сравнению с представителями других классов, обладающими всего одной слуховой косточкой. Предельные возможности слуха млекопитающих находятся в диапазоне от 20—30 Гц до 150—200 кГц, т. е. простираются далеко в область ультразвуковых частот. Слуховые центры позвоночных. Необходимость восприятия малых акустических уровней, развиваемых звуковой волной, определила возникновение в филогенезе позвоночных не только специализированных приемников, но и прогрессивное развитие мозговых слуховых центров. В связи с тем что волосковые клетки позвоночных, будучи вторичночувствующими рецепторами, лишены аксона, слуховые сигналы передаются в ЦНС нейроном второго порядка. Это биполярная клетка, тело которой находится в улитке. Периферическое волокно этой клетки образует синапсы с волосковыми клетками, тогда как центральный отросток входит в состав слухового нерва. Аксоны нейронов слухового нерва заканчиваются на телах и дендритах нервных клеток продолговатого мозга. У крутлоротых и хрящевых рыб первое центральное переключение слуховой импульсации находится в трех группах ядер продолговатого мозга: дорсальном, медиальном и вентральном. Считают, что вентральное ядро преимущественно связано с VIII парой черепных нервов, а дорсальное и медиальное — с системой органов боковой линии. Часть волокон идет также к обеим половинам мозжечка и к крупным двигательным клеткам ствола мозга. У костистых рыб распределение волокон акустико—латеральной системы отличается от строения этой системы у круглоротых и хрящевых рыб. Основными акустическими ядрами у них считают тангенциальное ядро и ядро Дейтерса. Часть волокон VIII нерва, в особенности саккулярные волокна, заканчиваются на дендритах маутнеровских клеток. Известно, что аксоны маутнеровских клеток иннервируют мускулатуру туловища рыбы. Аксоны клеток ядер продолговатого мозга идут в средний мозг и заканчиваются в высшем слуховом и оптическом центре, а также в ядре покрышки среднего мозга. Клетки центров среднего мозга имеют друг с другом двусторонние связи и являются высшими интегративными центрами звукового, зрительного и тактильного чувств. От этих центров берут начало пути, обеспечивающие сложные двигательные реакции животных. Переход от водной к воздушной среде обитания привел не только к появлению специализированных рецепторных структур, но и к выделению чисто слуховых волокон, проходящих в составе преддверно—улиткового нерва. Четко идентифицированные слуховые волокна определяются только у лягушек, большую часть жизни проводящих на суше, тогда как водные амфибии имеют лишь незначительную слуховую часть VIII нерва. Все слуховые волокна от амфибиальной и базилярной папилл оканчиваются в дорсальном ядре продолговатого мозга, которое можно считать гомологом улиткового ядра млекопитающих. Волокна от этих ядер идут к верхней оливе в составе пучка, представляющего собой зачаток трапециевидного тела, а от верхней оливы к среднему мозгу — в виде пучка, который в ходе эволюционного развития превращается в мощную систему латеральной петли. Четких ядерных групп в латеральной петле не обнаруживается. Слуховые пути, идущие от слуховых центров среднего мозга в передние отделы мозга, немногочисленны. Таким образом, улитковое ядро, трапециевидное тело, верхняя олива, латеральная петля являются самыми древними центральными образованиями в эволюции слуховой системы у наземных позвоночных. В дальнейшем происходит увеличение объема и дифференциация этих древних отделов, а также проникновение слуховых волокон в передний мозг (таламус, конечный мозг, корковые структуры). После возникновения таламической и корковой проекции развитие идет по пути усложнения структурно—функциональной организации всех отделов слуховой системы. Рассмотрим это положение на примере млекопитающих. Схема восходящих слуховых путей у человека представлена на рис. 4.31.
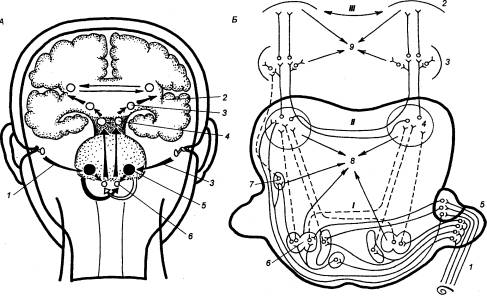
Существует пять основных уровней переключения восходящих слуховых волокон: улитковые ядра, верхнеоливарный комплекс, нижние холмики, медиальное коленчатое тело и слуховая зона коры больших полушарий (рис. 4.32). Кроме того, по ходу слухового пути расположено большое количество небольших ядер, в которых осуществляется частичное переключение восходящих слуховых волокон. У млекопитающих происходит разделение улиткового ядра на три основные части: передний и задний отделы вентрального улиткового ядра и дорсальное улитковое ядро, резко различающиеся по клеточному составу и системе связей. Каждое слуховое нервное волокно, войдя в ствол мозга и достигнув улитковых ядер делится на множество концевых ветвей. В ядре эти ветви распределены строго упорядоченным образом. Кроме того, вся тонотопическая последовательность рецепторов базилярной пластинки проецируется через слуховые волокна на различные участки улиткового ядра. Таким образом, одна улитка имеет в улитковых ядрах множественное представительство. Такой же процесс дифференцировки и распределения на несколько ядер касается верхней оливы, которая у современных высокоорганизованных млекопитающих состоит из группы ядер, входящих в состав верхнеоливного комплекса. Иннервация верхнеоливарного комплекса также тонотопическая. Ламинарная организация важнейших слуховых центров — нижних холмиков и медиального коленчатого тела — тоже предполагает тонотопическую организацию этих ядер, показанную в ряде электрофизиологических исследований. Это является одним из выражений принципа пространственного анализа звуков и одним из оснований тонкого частотного различения. Главным таламическим релейным ядром для слуховой информации является медиальное коленчатое тело. У высокоорганизованных млекопитающих область проекции релейных клеток медиального коленчатого тела на кору соответствует первичной слуховой коре. Традиционно считали, что в коре существует только одна первичная слуховая зона, однако новые работы выявили множественные области и в таламическом ядре, и в коре, а также соединяющие их параллельные пути. У многих млекопитающих в первичной коре находится область с крупной и подробной картой частот, называемая AI, что свидетельствует о тонотопическом принципе организации и коры больших полушарий. Многочисленными исследованиями показано существование в слуховой сенсорной системе нисходящих путей от височной коры к медиальному коленчатому телу, нижним холмикам, ядрам латеральной петли, верхней оливе, ядрам трапециевидного тела (см. рис. 4.32). Они начинаются в основном от первичного слухового поля коры AI и идут вместе с восходящими. Окончания кортико—фугальных волокон от височной области коры были обнаружены и в других неспецифических слуховых образованиях головного мозга.
|