В домене Bacteria выделено более 15 основных эволюционных линий
Число признанных родов архей с 1977 г. возросло от 5 до примерно 40, тогда как число родов бактерий почти не изменилось. Вместе с родами цианобактерий, которые могут быть описаны согласно Международному кодексу номенклатуры бактерий и Международному кодексу ботанической номенклатуры, это число равно примерно 400.В противоположность Archaea, для которых фактически определено филогенетическое положение каждого типового штамма, ближайшее филогенетическое соседство более чем 80 родов бактерий к настоящему времени не установлено. Выделяют примерно 20 четко разграниченных отдельных эволюционных линий; однако большинство видов бактерий относится к двум из них — грамположительным бактериям и Proteobacteria. В табл. 2 сопоставлены высшие таксоны домена Bacteria и указано приблизительное число родов в каждом из них. Поскольку студентам важно в первую очередь ознакомиться с основными революционными изменениями в систематике прокариот, не вдаваясь в мелкие детали родственных связей, мы не будем касаться здесь родственных связей на родовом и видовом уровнях. Многие роды до сих пор представляют собой филогенетически гетерогенные таксоны (например, Rhodopseudomonas, Pseudomonas, Blasthobacter и Thiobacillus среди Proteobacteria и Bacillus, Clostridium, Desulfotomaculum и Peptococcus среди грамположительных бактерий).
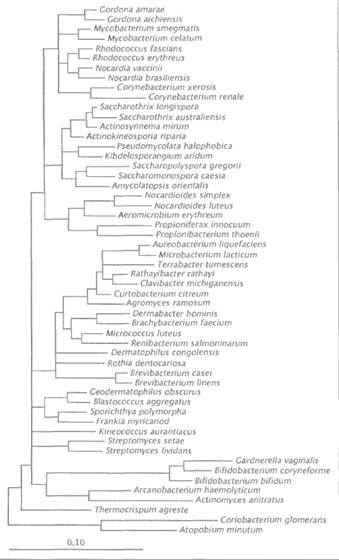
Таблица 2. Основные филогенетические линии внутри домена Bacteria по результатам анализа 16S-рРНК. В скобках указано число родов отнесенных к каждой линии (за исключением цианобактерий) Линия Aquifex (3) Термофильные аэробы (6) Линия Deinococcus (3) Линия Thermodesulfobacterium (1) Линия Bacteroides/Cytophaga (24) Линия Chloroflexus (6) Линия спирохет (10) Линия Leptospirillum (3) Линия Phusobacterium (4) Линия хламидий (1) Линия зеленых серных бактерий (5) Линия Planctomycetales (4) Линия Verrucomicrobiales (1) Линия Synergistes (1) Линия Acidobacterium (1) Линия Phibrobacter (1) Линия протеобактерий (~270) Линия грамположительных бактерий (~172) Линия цианобактерий Как пример успехов таксономии можно рассматривать порядок Actinomycetales, который в настоящее время представляет собой филогенетически хорошо очерченный таксон. Проанализированы последовательности почти каждого типового штамма типовых видов, и топология филогенетических деревьев этого порядка может считаться стабильной (рис. 5). Четыре эволюционные линии бактерий — грамположительные бактерии, Proteobacteria, таксон Bacteroides / Cytophaga / Flavobacterium и Cyanobacteria — имеют сложную филогенетическую структуру. Другие линии (например, включающие роды Thermotoga, Chloroflexus, Thermomicrobium и Herpetosiphon, Deinococcus и Thermus, Chlorobium, Chloroherpeton и Clathrochloris, а также Spirochaetales, Planctomycetales и некоторые другие филогенетические линии; табл. 2), определены на основе анализа лишь нескольких видов или даже одного единственного вида. Заключения широкого плана в области филогении, касающиеся филогенетических связей на уровнях от домена до семейств, основаны главным образом на результатах анализа 16S-рРНК. Только родственные связи представителей грамположительных бактерий, Proteobacteria и филогенетической линии Cytophaga были проанализированы несколькими различными методами, такими как гибридизация ДНК—ДНК, определение сходства цистронов рРНК и анализ последовательностей 23S-рРНК.
Рис. 5. Филогенетическая структура порядка Actinomycetales. Показаны взаимосвязи между семействами и внутри семейств, определенные путем анализа матрицы расстояний. Шкала — 10% дивергенция последовательностей. В результате этих исследований было установлено, что несколько традиционно признаваемых таксонов представляют собой также и филогенетически однородные группы, например актиномицетов (см. рис. 5), спирохет (на основании морфологии), миксобактерий, составляющих единый кластер в пределах сигма-подкласса протеобактерий (по признаку общественного поведения), и цианобактерий (по признаку фотосинтеза с выделением кислорода). Термофилия, фототрофия, механизм подвижности и сложная морфология (спиральная форма, почкование, простеки, мицелий) в основном могут рассматриваться как малоинформативные таксономические маркеры для определения рангов на высших уровнях, поскольку каждая из этих особенностей встречается независимо у объектов, находящихся в разных местах на филогенетическом дереве. Линия грамположительных бактерий разделяется на две основные ветви, при этом каждая из ветвей содержит глубоко укореняющиеся организмы, которые отрицательно окрашиваются по Граму. Грамотрицательный род Fusobacterium и близкие к нему роды, возможно, связаны с грамположительными организмами, но вопрос о том, составляют ли грамположительные бактерии филогенетически связанную группу, пока остается спорным. Разделение их на две основные ветви коррелирует с характерными для их представителей величинами содержания G + С в ДНК (ветвь клостридий, низкое содержание G + С, и ветвь актиномицетов, высокое содержание G + С), хотя установлены некоторые исключения из этого общего правила в обеих ветвях. Порядок ветвления филогенетического дерева таксонов грамположительных бактерий, основанный на результатах анализа 16S-рРНК, чрезвычайно информативен для таксономических выводов и ревизий, поскольку в большинстве случаев позволяет выявлять группы действительно родственных видов путем сопоставления хемотаксономических и других фенетических признаков. Такие характеристики, как состав клеточных стенок, структура липидов, жирных кислот, полиаминов и изопреноидных хинонов, нуклеотидный состав ДНК, отношение к кислороду и некоторые другие, являются информативными признаками грамположительных бактерий, позволяющими проводить четкое разграничение таксонов по сочетанию химических, физиологических и морфологических особенностей. Признаки, традиционно используемые в качестве единственного критерия для описания высших таксонов, такие как образование спор и морфологические особенности, теряют значимость в филогенетической таксономии, поскольку имеют полифилетическое происхождение или могли быть утрачены в ходе эволюции. Для класса Proteobacteria характерна сходная, однако значительно более сложная ситуация. В нем на основе филогенетического анализа выделены подклассы, обозначенные буквами греческого алфавита — от альфа до эпсилон. В этом классе идентифицировано более 220 родов, большинство из которых входит в альфа-подкласс (>70 родов) и гамма-подкласс (>90 родов). Едва ли хотя бы одно классическое таксономическое свойство, считавшееся пригодным для выявления естественных родственных связей, такое как фототрофия, скользящее движение, размножение почкованием, автотрофия, литотрофия или местообитание, позволяет определять здесь важную эволюционную ветвь. В пределах коротких филогенетических расстояний можно отметить совершенно различные фенотипы (фототрофы и СО-окисляющие организмы, фототрофы и литотрофы, азотфиксаторы и фитопатогенные бактерии); организмы, образующие ассоциации с высшими организмами, более родственны между собой, чем считалось ранее (например, Agrobacterium, Rhizobium, Brucella и Rochalimaea в альфа-подклассе). Удивителен состав сигма-подкласса, который не может быть объяснен на основании какого-либо другого молекулярного свойства, поскольку в него входят чрезвычайно сложноорганизованные миксобактерии, род Bdellovibrio, характеризующийся хищничеством в отношениии других бактерий, и анаэробные грамотрицательные бактерии, восстанавливающие сульфат и серу, такие как Desulfovibrio и Desulfuromonas.
Специфические родственные связи обнаружились неожиданно между фенотипически не сходными таксонами, такими как грамположительные, устойчивые к радиации деинобактерии и Грамотрицательный род Thermus, а также между строго анаэробным родом Bacteroides и эволюционными линиями аэробов Cytophaga и Flavobacterium. Поскольку представители каждой из этих двух фенотипически весьма разнородных пар организмов никогда не рассматривались как родственные, их ранее никогда не включали в совместный фенотипический анализ. Данных в пользу такого группирования пока также немного, но его достоверность подтверждается тем, что и деинобактерии, и виды Thermus содержат в составе пептидогликана остатки лизина и в случае линии Bacteroides-Cytophaga общим характерным признаком является наличие сфинголипидов. Рис. 6. Филогенетическое положение некоторых представителей родов альфа-, бета-, и гамма-подклассов класса Proteobacteria. Шкала — 10% дивергенция поспедовательностей.
Примеры того, что какие-либо организмы филогенетически неродственны между собой, несмотря на наличие общего и явно филогенетически значимого признака, встречаются редко. Наиболее яркий пример такого рода — отсутствие родства между родами Chloroflexus и Chlorobium. Оба эти таксона характеризуются наличием в клетках хлоросом, сходных по структуре и функции, но при этом содержат реакционные центры различных типов. Возможно ли, что такая сложная морфологическая структура возникла независимо в неродственных группах, подвергались ли гены, связанные с синтезом хлоросом, горизонтальному переносу, достоверны ли заключения на основе анализа 16S-рРНК — на все эти вопросы должны дать ответ дальнейшие исследования. Новый ключ к их решению этих вопросов дает открытие того, что родственные Chloroflexus роды, такие как Thermomicrobium и Herpetosiphon, лишены хлоросом, тогда как роды, родственные Chlorobium (такие как Chloroherpeton и Clathrochloris), содержат хлоросомы.
|