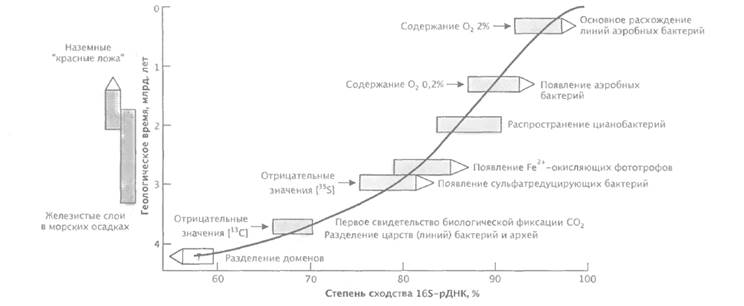
Палеохимические и геологические данные некоторым образом коррелируют с порядком ветвления, установленным по данным 16S-рРНК-анализа
Ископаемые находки в осадочных скалах докембрийского периода весьма скудны. Древнейшие ископаемые биологические объекты известны из осадков Исуа (Гренландия) возрастом 3,8 млрд. лет, группы Уааравуны (Австралия) возрастом 3,5 млрд. лет и формации Сваркоппи (Южная Африка). Несмотря на то, что ископаемые находки не позволяют сколько-нибудь основательно выяснять биохимическую природу древних объектов, они все же свидетельствуют, что жизнь существовала уже 3,2 млрд. лет назад. Данные палеохимии Более информативен, по сравнению с анализом ископаемых находок, химический анализ неорганических соединений в составе геологически древних осадочных пород (рис. 7). Увеличение соотношения [12С]орг./[13С]орг. обычно рассматривается как показатель фиксации углерода при фотосинтезе или других автотрофных процессах фиксации углерода. В биологических остатках содержание более легкого изотопа [12С]орг. выше по сравнению с изотопом [13С]орг.
Рис. 7. Последовательность появления основных групп бактерий в процессе эволюции. Фактор дискриминации сигма для [13С]орг. составляет 25‰. Эта величина, которая была установлена также в материале из породы возрастом 3,8 млрд. лет в осадках Исуа, оставалась относительно постоянной на протяжении миллиардов лет, т. е. скорость продукции биомассы фактически не изменялась за время биологической эволюции. Распространение механизмов фиксации СО2 у представителей обоих доменов наводит на мысль, что эволюция гипотетического предка — прогенота — по двум направлениям эволюционных линий прокариот происходила за довольно «короткий» период (первые 700 млн. лет) после образования планеты Земля. Не исключено, что разделение изотопов углерода имело место даже ранее, чем 3,8 млрд. лет назад, но их не удается определить в осадочных породах. Первым источником энергии и восстановителя для фиксации СО2 предположительно служило образование пирита. По этой гипотезе, высказанной Г. Вехтерхойзером, Н2S играет важную роль как предшественник молекул с сульфгидрильными группами в каталитических центрах, тогда как пириты рассматриваются в качестве предшественников железосерных центров и ферредоксина. Возможно, к предковым биохимическим типам относятся термофильные и хемолитоавтотрофные представители Crenarchaeota, на что указывает их биохимический потенциал. Важные палеохимические данные дает анализ отношения [32S]/[34S], величина которого возрастает в результате биологического восстановления сульфата. В осадочных породах Исуа, имеющих возраст 3,8 млрд. лет, такого возрастания не обнаружено. Наиболее раннее заметное изменение этого отношения выявлено в породах, имеющих возраст 3,2-2,8 млрд. лет. Сульфатное дыхание, известное у Desulfotomaculum (грамположительная бактерия) и Desulfovibrio, а также у родственных бактерий (Proteobacteria), может быть, таким образом, более поздней адаптацией, чем фотосинтез и метаногенез. Это предположение согласуется с результатами анализа 16S-рРНК, которые позволяют разделить эти линии сульфатредукторов примерно на уровне 78% сходства. Главное расхождение линий грамотрицательных сульфатредукторов, возможно, произошло на еще более поздней стадии эволюции (примерно 82% сходство). Аналогичным образом можно анализировать историю появления в атмосфере молекулярного кислорода, события которой важны для примерного датирования более поздних периодов эволюции с использованием анализа сходства последовательностей. Почти не вызывает сомнений, что до появления растительного фотосинтеза свободный кислород присутствовал в атмосфере не более чем в следовом количестве. Источник кислорода в раннем докембрии неизвестен; маловероятно, что им мог быть фотолиз водяного пара в верхних слоях атмосферы. Существование железистого слоя в составе морских донных отложений (возрастом примерно 3,2-1,9 млрд. лет), который содержит магнетит и гематит, может объясняться жизнедеятельностью оксигенных прокариот. Остатки организмов, напоминающих цианобактерии, присутствуют в строматолитах возрастом 2,7 млрд. лет, но их таксономическое положение пока не установлено; главное расхождение линий этих организмов происходило позднее, чем 3,2-2,7 млрд. лет назад. Если, тем не менее, образование железистых слоев в морских донных отложениях вызывала жизнедеятельность биологических объектов, то либо кислород должны были выделять организмы, для которых не описано потомков, либо окисление Fе2+ до Fе3+ осуществлялось без участия кислорода анаэробными фототрофами. Более поздние типы фототрофов, родственные современным фототрофам, как недавно установлено, обладают такой метаболической активностью. После окисления в морской среде всех доступных для этого соединений началось формирование наземных железистых образований, известных под названием красные ложа (возраст 2,0-1,8 млрд. лет). Их существование можно объяснять активностью цианобактерий, так как в строматолитах формации Ганфлит, имеющей возраст 2,0 млрд. лет, обнаружено большое разнообразие остатков цианобактерий. Интересно, что отмечается корреляция между возрастанием уровня кислорода 1,5-1,9 млрд. лет назад и появлением микроаэрофильных организмов Lactobacillus, Streptococcus, Actinomyces, спирохет и некоторых представителей Proteobacteria, которые эволюционировали от анаэробных фототрофных и гетеротрофных предков в различных филогенетических линиях бактерий. Только после того как была достигнута точка Пастера (0,2% O2; 1,2-1,4 млрд. лет назад), для организмов стало предпочтительным переключение с брожения на аэробное дыхание (рис. 29.31). Разделение ветвей аэробных форм, эволюционировавших от микроаэрофильных предковых организмов-бродилыциков, коррелирует со сходством рРНК приблизительно на уровне 87%, тогда как основное расхождение строго аэробных организмов имело место при значениях сходства выше 92%. Это событие могло произойти при парциальном давлении кислорода 2%, в нижнем силуре (примерно 440 млн. лет назад).
|