Фоторецепция
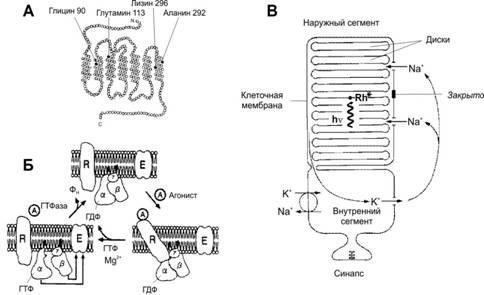
В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин (рис. 10–5А) состоит из белковой части (опсин) и хромофора — 11- цис -ретиналя, под действием фотонов переходящего в транс -ретиналь (фотоизомеризация). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события (рис. 10–5Б): активация родопсина в результате фотоизомеризации ® каталитическая активация G‑белка (Gt, трансдуцин) родопсином ® активация фосфодиэстеразы при связывании с Gta ® гидролиз цГМФ цГМФ–фосфодиэстеразой ® переход цГМФ–зависимых Na+‑каналов из открытого состояния в закрытое ® гиперполяризация плазмолеммы фоторецепторной клетки ® передача сигнала на биполярные клетки.
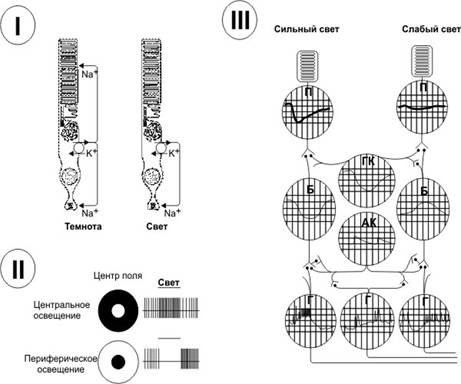
Рис. 10 – 5. РОДОПСИН И АКТИВАЦИЯ ИОННЫХ КАНАЛОВ [11]. А. Молекула опсина содержит 7 трансмембранных альфа-спиральных участков. Зачернённые кружки соответствуют локализации наиболее распространённых молекулярных дефектов. Так, при одной из мутаций глицин во втором трансмембранном участке в 90-м положении заменён на аспарагин, что приводит к врождённой ночной слепоте. Б. Трансмембранный белок родопсин и его связь с G‑белком (трансдуцин) в плазмолемме фоторецепторной клетки. Возбуждённый фотонами родопсин активирует G‑белок. При этом гуанозиндифосфат, связанный с a‑СЕ G‑белка, заменяется на ГТФ. Отщеплённые a‑СЕ и b‑СЕ действуют на фосфодиэстеразу и заставляют её превращать цГМФ в гуанозинмонофосфат. Это закрывает Na+‑каналы, и ионы Na+ не могут попасть в клетку, что приводит к её гиперполяризации. R — родопсин; a, b и g — СЕ G‑белка; A — агонист (в данном случае кванты света); E — фермент–эффектор фосфодиэстераза. В. Схема палочки. В наружном сегменте расположена стопка дисков, содержащих зрительный пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет (hn) активирует родопсин (Rh*) в дисках, что закрывает Na+‑каналы в клеточной мембране и снижает вход Na+ в клетку. · Ионные основы фоторецепторных потенциалов à В темноте Na+‑каналы мембраны наружных сегментов палочек и колбочек открыты, и ток течёт из цитоплазмы внутренних сегментов в мембраны наружных сегментов (рис. 10–5В и 10–6I). Ток течёт также в синаптическое окончание фоторецептора, вызывая постоянное выделение нейромедиатора. Na+,K+–насос, находящийся во внутреннем сегменте, поддерживает ионное равновесие, компенсируя выход Na+ входом K+. Таким образом, в темноте ионные каналы поддерживаются в открытом состоянии и потоки внутрь клетки Na+ и Ca2+ через открытые каналы обеспечивают появление тока (темновой ток).
Рис. 10 – 6. ЭЛЕКТРИЧЕСКИЕ РЕАКЦИИ СЕТЧАТКИ. I. Ответ фоторецептора на освещение. II. Ответы ганглиозных клеток. Освещённые поля показаны белым. III. Локальные потенциалы клеток сетчатки. П — палочки, ГК — горизонтальные клетки, Б — биполярные клетки, AК — амакринные клетки, Г — ганглиозные клетки. à На свету, т.е. когда свет возбуждает наружный сегмент, Na+‑каналы закрываются и возникает гиперполяризационный рецепторный потенциал. Этот потенциал, появившийся на мембране наружного сегмента, распространяется до синаптического окончания фоторецептора и уменьшает выделение синаптического медиатора — глутамата. Это немедленно приводит к появлению ПД в аксонах ганглиозных клетках. Таким образом, гиперполяризация плазмолеммы — следствие закрытия ионных каналов. Вышеописанный каскад реакций усиливает световой сигнал и объясняет удивительный факт, каким образом один фотон света — наименьшая возможная квантовая единица световой энергии — может вызывать измеряемый рецепторный потенциал величиной в 1 мВ, а 30 фотонов света могут повышать функциональную готовность палочкового аппарата сетчатки на 50%. à Возврат к исходному состоянию. Свет, вызывающий каскад реакций, понижающих концентрацию внутриклеточного цГМФ и приводящих к закрытию натриевых каналов, уменьшает содержание в фоторецепторе не только Na+, но и Ca2+. В результате понижения концентрации Ca2+ активируется фермент гуанилатциклаза, синтезирующая цГМФ, и в клетке растёт содержание цГМФ. Это приводит к торможению функций активированной светом фосфодиэстеразы. Оба этих процесса — повышение содержания цГМФ и торможение активности фосфодиэстеразы — возвращают фоторецептор в исходное состояние и открывают Na+‑каналы. · Световая и темновая адаптация à Световая адаптация. Если человек длительное время находится в условиях яркого освещения, то в палочках и колбочках происходит превращение значительной части зрительных пигментов в ретиналь и опсин. Большая часть ретиналя превращается в витамин A. Всё это приводит к соответствующему снижению чувствительности глаза, называемому световой адаптацией. à Темновая адаптация. Напротив, если человек остаётся в темноте продолжительное время, то витамин A вновь превращается в ретиналь, ретиналь и опсин формируют зрительные пигменты. Всё это приводит к повышению чувствительности глаза — темновой адаптации. Ú При нахождении в темноте световая чувствительность палочек нарастает неравномерно: в первые минуты она увеличивается в десятки раз, в конце первого часа чувствительность палочек к свету возрастает до сотен тысяч раз. В темноте пигменты колбочек восстанавливаются быстрее, чем родопсин палочек, но это не приводит к заметному повышению чувствительности глаза, поскольку абсолютная чувствительность колбочек к свету незначительна. Ú В дополнение к адаптации, вызываемой изменениями концентрации родопсина и зрительных пигментов, глаз имеет и другие механизмы адаптации. Так, изменение размеров зрачка в течение долей секунды может в 30 раз уменьшить поступление света к сетчатке. Ú Другой механизм связан с функциональными изменениями нейронов сетчатки. В темноте увеличивается число возбуждённых ганглиозных клеток, что приводит к возрастанию световой чувствительности. ЦНС также влияет на адаптацию сетчатки к действию света. Так, засветка одного глаза понижает чувствительность неосвещённого глаза. Световая чувствительность глаза может изменяться и под воздействием звука.
электрические ответы сетчатки Различные клетки сетчатки (фоторецепторы, биполярные, горизонтальные, амакринные, а также дендритная зона ганглиозных нейронов) генерируют локальные потенциалы, но не ПД (рис. 10–6). Из всех клеток сетчатки ПД возникают только в аксонах ганглиозных клеток. Суммарные электрические потенциалы сетчатки — электроретинограмма (ЭРГ). ЭРГ регистрируют так: один электрод накладывают на поверхность роговицы, другой — на кожу лица. ЭРГ имеет несколько волн, связанных с возбуждением различных структур сетчатки и суммарно отражает интенсивность и длительность действия света. Данные ЭРГ могут использоваться в диагностических целях при заболеваниях сетчатки · Связи между клетками сетчатки à Фоторецепторы образуют синапсы с биполярными нейронами. В области центральной ямки расположены преимущественно колбочки. Каждая колбочка центральной ямки образует синапс только с одним биполярным нейроном. à Биполярные нейроны синаптически связаны с ганглиозными и амакринными клетками. Популярна концепция о том, что ограниченное число биполярных клеток передает информацию 16 типам ганглиозных клеток при участии не менее 20 типов амакринных клеток. à Горизонтальные клетки формируют синаптические контакты с отростками фоторецепторов. Горизонтальные клетки получают информацию от колбочек и передают её также колбочкам. Соседние горизонтальные клетки связаны между собой щелевыми контактами. à Амакринные клетки. Их перикарионы находятся во внутренней части внутреннего ядерного слоя в области синапсов между биполярными и ганглиозными клетками. Эти клетки получают информацию от биполярных нейронов и передают её ганглиозным нейронам. Функции десятков подтипов амакринных клеток полностью не выяснены. Часть клеток участвует в формировании пути палочкового зрения в сетчатке, другие отвечают за начало и окончание световых сигналов, третьи следят за правильностью движения светового пятна. В целом амакринные клетки выполняют функцию вставочных нейронов, осуществляющих первоначальный анализ световых сигналов. à Ганглиозные клетки — крупные мультиполярные нейроны многих разновидностей. Биполярные и амакринные клетки образуют синапсы на ганглиозных нейронах. Их аксоны образуют зрительный нерв. à Мюллеровские клетки. Кроме нейронов, сетчатка содержит крупные клетки радиальной глии — мюллеровские клетки. Их ядра расположены на уровне центральной части внутреннего ядерного слоя. Наружные отростки заканчиваются микроворсинками, образуя наружный пограничный слой. Внутренние отростки имеют расширение (ножку) во внутреннем пограничном слое на границе со стекловидным телом. Глиальные клетки играют важную роль в регуляции ионного гомеостаза сетчатки. В частности, они снижают концентрацию K+ во внеклеточном пространстве, где концентрация этих ионов при световом раздражении резко увеличивается. Плазматическая мембрана мюллеровских клеток в области ножки характеризуется высокой проницаемостью для ионов K+. Мюллеровская клетка захватывает K+ из наружных слоёв сетчатки и направляет поток этих ионов через свою ножку в жидкость стекловидного тела. · Нейромедиаторы. Нейроны сетчатки синтезируют ацетилхолин, дофамин, L -глутаминовую кислоту, глицин, g-аминомасляную кислоту. Некоторые нейроны содержат серотонин, его аналоги (индоламины) и нейропептиды. Палочки и колбочки в синапсах с биполярными клетками секретируют глутамат. Разные амакринные клетки выделяют ГАМК, глицин, дофамин, ацетилхолин и индоламин, оказывающие тормозные эффекты. Нейромедиаторы для биполярных и горизонтальных не идентифицированы. Локальные потенциалы. Ответы палочек, колбочек и горизонтальных клеток являются гиперполяризующими (рис. 10–6,II), ответы биполярных клеток либо гиперполяризующие, либо деполяризующие. Амакринные клетки создают деполяризующие потенциалы. · Локальные потенциалы колбочек и палочек. Рецепторные потенциалы колбочек и палочек возникают одинаково быстро, но скорость завершения рецепторного потенциала палочек медленнее. Кривая соотношения амплитуды рецепторного потенциала к интенсивности светового стимула имеет одинаковую форму и для палочек, и для колбочек, но палочки во много раз чувствительнее. Ответная реакция палочек пропорциональна интенсивности стимулов на уровне освещения, который ниже порога для колбочек. С другой стороны, колбочки отвечают пропорционально интенсивности стимулов при таком высоком уровне освещения, когда ответы палочек максимальны и не могут больше изменяться. Различия порогов восприятия освещённости колбочками и палочками расширяет диапазон и устойчивость восприятия светового раздражения сетчаткой. Благодаря чрезвычайно низкому порогу восприятия палочки являются детекторами абсолютно малой освещённости, а колбочки реагируют на изменения освещённости в тех пределах, когда палочки уже достигли своего максимума. · Проведение сигналов от палочек и колбочек. Зрительные пути от палочек и колбочек имеют некоторые отличия. Нейроны и аксоны ганглиозных клеток, проводящие зрительные сигналы от колбочек, значительно толще, чем те, что проводят сигналы от палочек. Скорость проведения сигналов для колбочкового зрения в два раза выше, чем для проведения сигналов палочкового зрения. Для колбочек, эволюционно представляющих молодую и быструю систему проведения, нейронный путь имеет три звена: колбочки ® биполярные клетки ® ганглиозные нейроны. Более древняя палочковая система зрения имеет зрительный путь из 4 звеньев: палочки ® биполярные клетки ® амакринные клетки ® ганглиозные клетки.
|