Глава 1. Эвтрофирование озер и водохранилищ 2 страница
Специальное внимание в исследованиях влияния соотношения азота и фосфора уделялось изучению появления, развития и доминирования группы синезеленых водорослей в озерах. Физиологические особенности этой группы водорослей отличаются способностью фиксировать азот атмосферы, но даже не фиксирующие азот виды, например Microcystis aeruginosa, обладают способностью накапливать азот нитратов в клетках. Многолетние экспериментальные исследования удобряемых биогенами озер, проведенное Д.Шиндлером, убедительно подтвердило зависимость обилия цианобактерий от низких отношений азота к фосфору [Schindler, 1974]. Анализируя данные о видовом составе водорослей в 17 озерах, В.Смит пришел к выводу, что явное доминирование синезеленых водорослей начинается при отношении N:P менее 29:1, хотя получить достаточно четкую эмпирическую зависимость ему не удалось [Smith, 1982]. При количественном описании и моделировании процесса развития водорослей в водоемах проявились различные точки зрения на принципы биогенного лимитирования развития фитопланктона. В биологии доминирует принцип лимитирования Либиха, согласно которому скорость развития биологического процесса определяется тем элементом, концентрации которого находятся в наибольшем относительном дефиците. Однако закон Либиха применим только для стационарного сбалансированного состояния экосистемы. В периоды резких колебаний баланса энергии лимитирующие организмы могут быстро сменять друг друга. Поэтому многие специалисты по экологическому моделированию считают, что к экосистемам необходимо применять закон мультипликативного лимитирования, когда несколько элементов должны рассматриваться лимитирующими и, соответственно, необходимо учитывать концентрации всех этих элементов [Крышев, Сазыкина, 1990]. Результаты современных исследований показывают преимущество мультипликативных моделей лимитирования, воспроизводящих многофакторную зависимость развития фитопланктона от условий биогенного питания, хотя в моделировании биологических процессов в целях упрощения часто продолжают использоваться модели, построенные на принципе Либиха [Jorgensen, 1994].
1.4. Показатели трофического состояния водоемов
Принципиальный момент при оценке процесса эвтрофирования - определение трофического состояния водоемов, которое фактически отражает метаболизм экосистемы (поступление, накопление и расходование энергии). Основы типологической классификации трофического состояния водоемов были заложены еще классиками современной лимнологии Науманом и Тинеманом. [Thienemann, 1925, Naumann, 1932]. Предложенное Науманом разделение водоемов на олиготрофные, мезотрофные и эвтрофные до сих пор остается основополагающим при оценке развития водных экосистем. В действительности процесс эвтрофирования водоемов представляется (даже при ускоренном антропогенном эвтрофировании) непрерывно сглаженным от состояния, когда водная среда бедна биогенными веществами и малопродуктивна (олиготрофный водоем) до состояния избытка биогенных веществ и высокой продукции. Поэтому выделение дискретных состояний весьма условно и позволяет предложить лишь наиболее общие градации трофического состояния водоема. Иногда используют и дополнительные промежуточные категории – ультраолиготрофный, олиго-мезотрофный, мезо-эвтрофный, гиперэвтрофный водоем. Разделение водоемов по уровням трофии в настоящее время представляется одновременно и субъективным и относительным, поэтому в количественных оценках трофического состояния водоема немало сложностей и противоречий. Несмотря на то, что нет недостатка в предложениях классификаций и количественных индексов, оценка уровня трофии конкретного водоема остается весьма серьезной проблемой, актуальность которой усиливается тем, что адекватное количественное описание состояния водоема чрезвычайно важно при выборе стратегии охраны его экосистемы. Очевидно, что наиболее объективную оценку можно составить на основе полного лимнологического обследования водоема с возможно более детальным учетом пространственно-временной изменчивости характеристик экосистемы. Однако, общеизвестные трудности организации и проведения такого обследования вынудили искать более простые и однозначно связанные с процессом эвтрофирования показатели и критерии. В настоящее время при оценке трофического состояния водоема в лимнологии используется широкий набор биологических, химических и физических характеристик и показателей функционирования экосистемы. Наиболее распространенный и простой способ оценки трофического состояния водоема – использование классификаций на основе градаций показателей, характеризующих экосистему. Литература по этому вопросу чрезвычайно обширна. Исключительное разнообразие и изменчивость географических и экологических условий эвтрофирования обусловили появление большого количества различных классификаций, включающая перекрывающиеся границы трофических уровней.
1. Биологические и биохимические показатели. Поскольку сущность процесса эвтрофирования состоит в общем увеличении продуктивности экосистемы, в основе которого лежит рост первичной продукции, биологические показатели акцентированы на переменные, характеризующие процесс первичного продуцирования органического вещества. Первичная продукция водоемов. Содержание хлорофилла “a” в воде водоема.. Впервые систему разделения водоемов на классы по величине первичной продукции применил Г.Г.Винберг, которая была развита Г.Лайкенсом, привлекшим для классификации пресных вод данные по биомассе фитопланктона, содержанию хлорофилла “а” и общего органического углерода. Классификация Г.Лайкенса по этим показателям в дальнейшем была включена в классификацию трофических состояний OECD [Likens, 1975] Система градации озер северной и умеренной зоны детально разработана С.П.Китаевым по величинам годовой первичной продукции. [Китаев, 1984]. Здесь следует заметить, что годовую первичную продукцию нужно рассматривать как функцию ее суточных величин и длительности вегетационного сезона. Поэтому при одной и той же суточной продукции годовая продукция озер умеренных широт выше, чем северных и ниже, чем южных. Крупные южные озера олиготрофные по содержанию хлорофилла, скорости фотосинтеза на оптимальной глубине, могут быть мезотрофными по годовым величинам продукции. Для разработки трофической классификации, учитывающей особенности как “планктонных”, так и “детритных” озер, важно выбрать основополагающий критерий трофности. Следует подчеркнуть, что “планктонные” и “детритные” озера представляют собой два крайних типа экосистемы, различающихся по структуре трофической цепи. Для первого типа экосистем более вероятен поток энергии, направленный по пастбищной цепи, для второго типа - по детритной цепи с непременным участием гетеротрофных микроорганизмов. Однако, большинство экосистем относится к смешанному типу, так как пастбищная и детритная цепи в них тесно взаимосвязаны. Несмотря на очевидные преимущества использования для оценок трофности динамического показателя, а именно величины первичной продукции, по сравнению со статическим показателем – содержанием хлорофилла-а, последний гораздо чаще фигурирует в существующих системах классификации трофического состояния водоемов. Основной проблемой использования этого показателя представляется исключительно высокая его пространственно-временная изменчивость [Бульон, 1993]. Как показывает практика, при нерегулярных наблюдениях содержание хлорофилла “а” в планктоне точнее отражает трофический статус озер, чем скорость фотосинтеза, легко реагирующая на изменения погодных условий. Иными словами, содержание хлорофилла “а” - более консервативный показатель трофности и поэтому при соблюдении определенных условий полнее передает продукционный потенциал фитопланктона. Классификации трофического состояния водоемов по состоянию фитопланктона приведены в табл. 1.1.
Таблица 1. 1 Классификации типов водоемов по содержанию хлорофилла-а.
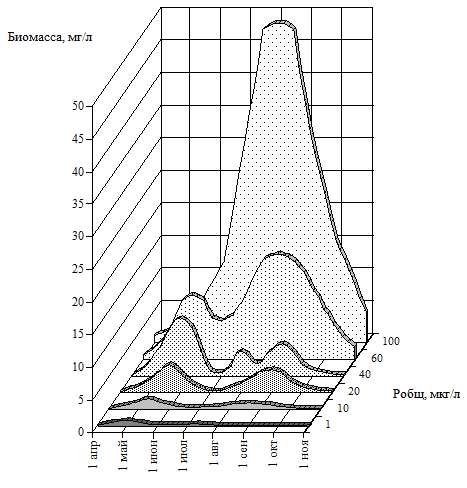
Распространенность этого показателя в качестве индикатора трофического состояния связана также с относительной простотой его определения и, следовательно, возможностью получения массовых материалов по водоемам, а также тесной функциональной связью с продукционными характеристиками экосистемы и биомассой фитопланктона. Обобщение многочисленных эмпирических связей между показателями трофности водоемов приведено в монографии А.П.Мусатова [Мусатов, 2001]. Серьезной проблемой выступает исключительная пространственно-временная изменчивость продукционных характеристик. Известная пятнистость цветения водоема, сильная зависимость продукционных характеристик от сгонно-нагонных явлений и погодных условий требует проведения учащенных наблюдений, анализа данных и соответствующего осреднения наблюдаемых значений. Однако использование лишь одного, пусть даже самого значимого показателя, особенно учитывая его большую изменчивость, существенно обедняет оценку трофности и при недостаточно надежном исходном материале может привести к ошибочным оценкам. Поэтому, наряду с показателями первичной продуктивности водоемов, в практике оценки эвтрофирования широкое распространение получили и другие показатели состояния экосистемы водоема. Биомасса фитопланктона. Учет биомассы фитопланктона стандартным методом микроскопирования, как известно, довольно трудоемок, поэтому надежный материал, характеризующий состояние водоема по этому показателю получить трудно. Этим объясняется сравнительно редкое использование биомассы фитопланктона при оценках эвтрофирования водоемов. Специальными гидробиологическими исследованиями показано, что содержание хлорофилла-а в единице биомассы (удельное содержание хлорофилла-а) зависит как от видового состава фитопланктона, так и от уровня биомассы фитопланктона, с увеличением которой оно убывает [Трифонова, Десортова, 1983, Елизарова, 1993]. Значительные сезонные колебания биомассы фитопланктона показывают, что наиболее надежной характеристикой для оценки трофического статуса представляются средние за сезон ее величины. Шкала трофности водоемов по средним за сезон величинам биомассы водорослей на основе лимнологического анализа озер предложена И.С.Трифоновой [Трифонова, 1990]. По мере эвтрофирования водоема размах отношения максимальных и минимальных величин биомассы фитопланктона увеличивается, что также может служить показателем трофического состояния при многолетних обследованиях водоема. Тесная (r=0.93) линейная связь между биомассой фитопланктона и степенью ее вариабельности установлена для Рыбинского водохранилища [Корнева, 1993]. Однако количественных оценок характерных величин изменчивости в водоемах разной трофии пока не предложено. Важной характеристикой, которая может использоваться при оценке трофического состояния водоема представляется характер сезонной динамики фитопланктона. Так, для олиготрофных водоемов характерно наличие одного небольшого максимума биомассы весной, для мезотрофных – раннелетней депрессии (на фоне умеренного развитии водорослей), а в эвтрофных водоемах на протяжении всего сезона может наблюдаться очень значительная биомасса фитопланктона (рис. 1.1).
Рис. 1.1. Схема изменения сезонной динамики биомассы фитопланктона в озерах умеренной зоны по мере усиления эвтрофирования (по: Трифонова, 1990).
И, наконец, спектр размеров клеток фитопланктона представляется весьма информативной характеристикой взаимодействия различных компонентов водных экосистем и их трофического состояния [Sprules, Munawar, 1986]. Классификации трофического состояния водоемов по биомассе фитопланктона представлены в табл. 1.2.
Таблица 1.2 Классификации типов водоемов по биомассе фитопланктона (мг/л)
Известная пятнистость цветения водоема, сильная зависимость продукционных характеристик от сгонно-нагонных явлений и погодных условий требует проведения учащенных наблюдений, глубокого анализа данных и соответствующего осреднения наблюдаемых значений. Значительные сезонные колебания биомассы фитопланктона показывают, что наиболее надежной характеристикой для оценки трофического статуса представляются средние за сезон ее величины. Детальный анализ пространственной неоднородности фитопланктона Рыбинского водохранилища, проведенный Корневой Л.Г с применением разнообразных статистических методов, показал, что применение классификаций по биомассе фитопланктона по данным гидробиологических съемок может привести к существенной разнице в оценках трофии различных участков водоема [Корнева, 1993]. Чтобы определить возможность использования данных мониторинга летнего фитопланктона для оценки трофического состояния водоема, необходимо отслеживать особенности пространственного распределения его биомассы по водоему. При биомассах, типичных для отдельно взятых водоемов и сезонов, коэффициенты корреляции между биомассой водорослей и содержанием хлорофилла-а оказываются высокими и, учитывая незначительность отмеченных отклонений, могут быть приняты прямолинейными, поэтому для ориентировочной характеристики трофического состояния водоемов вполне приемлемо использовать более распространенный показатель хлорофилл-а. Индикаторы трофического состояния по составу биологических сообществ. Впервые еще В.Пирселл указал на возможность индикации трофности озера по видовому составу фитопланктона [Pearsal, 1932]. Из многочисленных биологических показателей трофического состояния водоемов наибольшее распространение получили индикаторные виды организмов фитопланктона, зоопланктона и зообентоса. Изменение видов, которое происходит при эвтрофировании, носит вполне закономерный характер и может использоваться как характеристика процесса. Обычно считается, что начало доминирования синезеленых водорослей свидетельствует о прогрессирующем эвтрофировании водоема. Следует, однако, иметь в виду, что географические колебания видов являются слишком значительными, чтобы можно было подготовить их стандартный и общепринятый лимнологами перечень. Наиболее распространенный среди гидробиологов России список видов Зелинки-Марвана не раз подвергался справедливой критике и условность его применения очевидна, хотя для некоторых достаточно распространенных видов фитопланктона индикаторная оценка по этому списку вполне приемлема. Использовать видовой состав фитопланктона предлагалось различными авторами [Reynolds, 1998, Huszar et. аl., 1998]. Известен, например индекс Г.Нигарда, который для оценки трофического состояния озер применял соотношение различных групп водорослей [Nygaard, 1956]. Особенно успешным для оценки трофии может быть анализ фитопланктонных сообществ в тех случаях, когда более простые индикаторы либо дают противоречивые результаты, либо просто малоэффективны. Примером могут служить тропические водоемы, в которых как содержание хлорофилла-а, так и концентрация биогенных элементов недостаточно адекватны трофическому состоянию экосистемы вследствие высоких скоростей метаболических процессов. Детальное исследование структуры фитопланктонных сообществ генетически и морфологически различных водоемов Бразилии показало явную предпочтительность использования видового состава водорослей для оценки трофического состояния по сравнению с традиционными критериями ОЕРК [OECD…, 1982]. Помимо видового состава индикаторами трофии могут быть и некоторые физиологические характеристики фитопланктона, например, скорость роста. Стокнер, изучая отложения диатомей в озерах, разработал критерии определения трофической эволюции озер по соотношению Araphinidineae и Centrales [Stockner, 1975]. Диатомовый анализ состояния озер успешно применялся еще на заре лимнологических исследований, в частности, при анализе трофических изменений озер [Pearsall, 1932]. Однако, эти таксономические индексы не нашли широкого применения при оценке эвтрофирования на основе мониторинга. Статистический анализ показал их слабую корреляцию с традиционными методами оценки трофического состояния, например, по содержанию хлорофилла-а. [Миллиус, 1983]. И все же, как показали последующие исследования, индикаторные критерии эвтрофирования слабо коррелируют с полной комплексной оценкой трофического состояния по химическим и биохимическим показателям. Еще Дж. Хатчинсон критически оценивал возможность использования индикаторых организмов, обнаружив в одном озере одновременно типично олиготрофные и типично эвтрофные виды фитопланктона [Хатчинсон, 1963]. Видимо, здесь проявляется исключительная сложность и многофакторность процессов развития биологических сообществ, которая в отдельных индивидуальных экосистемах может затушевывать главные причинные механизмы процесса эвтрофирования. Зообентос считается одним из самых чувствительных компонентов экосистемы водоема к ее изменениям. Именно сообщества донных животных, в отличие от планктонных организмов, в силу присущей им инертности служат индикаторами изменений, происходящих в экосистеме водоема. В этом индексе находит отражение, как уменьшение разнообразия фауны, так и упрощение трофических связей по мере эвтрофирования, поэтому данные, полученные по известному методу Вудивисса, часто довольно объективно отражают реальную ситуацию в водоемах. Следует, однако, иметь в виду, что развитие донной фауны находится в сильной зависимости от кислородного режима гиполимниона водоема, поэтому бентос в некоторой степени опосредованно реагирует на изменение трофического статуса. Возможность использования структурно-функциональных характеристик зоопланктона для индикации эвтрофирования озерных экосистем была показана И.Н.Андронниковой [Андроникова, 1996]. Наиболее надежными и информативными показателями процесса эвтрофирования, по мнению автора, являются: - уменьшение числа доминирующих видов и снижение индекса видового разнообразия, - изменение процентного соотношения таксономических групп в сторону увеличения доли коловраток и кладоцер и уменьшения доли копепод, - изменение размерной структуры сообщества в сторону преобладания мелких форм и, как следствие, уменьшение среднего веса особи, - увеличение внутригодовой амплитуды колебаний биомассы, - снижение доли хищников в общей биомассе зоопланктона, - увеличение доли трат на обмен от биомассы зоопланктона, уменьшение соотношения биомасс зоопланктона и фитопланктона за вегетационный период. Классификации трофического состояния водоемов по зоопланктону приведены в табл. 1.3.
Таблица 1.3 Классификация трофического состояния водоемов по структурным показателям зоопланктона
В таблице: Q – отношение числа видов р.Brachionus к числу видов р.Trichocera E/O – соотношение числа видов-индикаторов эвтрофного и олиготрофного типа E = K(X+1)/(A+V)(Y+1), где K – число видов Rotatoria, A – Copepoda, V – Cladocera, X – мезотрофные виды, Y – олиготрофные виды Н = -S рilog2pi pi = Bi/B Bi – биомасса индикаторного вида зоопланктона, В – общая масса зоопланктона
Важно подчеркнуть, что значение имеет не столько величина этих показателей, сколько направленность их изменений в многолетнем ряду. Для выявления направленности многолетних изменений трофического статуса водоемов необходимы многолетние ряды наблюдений. Сравнивать показатели за отдельные годы можно только для лет со сходными гидрометеорологическими условиями (температура, инсоляция, водность, режим уровня), поскольку в годы с разными условиями возможны флуктуации, маскирующие тренд. Трофический статус водоемов нередко оценивается по состоянию фитопланктона. Действительно, между биогенной нагрузкой и количественным развитием фитопланктона (его продукцией, биомассой, содержанием хлорофилла в воде) для большинства исследованных озер и водохранилищ найдена прямая связь. Однако, связь эта нарушается в водоемах, сильно зарастающих погруженными макрофитами, в так называемых “макрофитных” экосистемах, где основными продуцентами являются высшие растения. В таких водоемах при возрастании биогенной нагрузки может не наблюдаться усиления развития фитопланктона и снижения прозрачности воды. Причина такого развития событий - связывание биогенов в тканях макрофитов, в которых кроме того может откладываться избыточный их запас, не используемый для дальнейшего продуцирования органического вещества ни самими макрофитами, ни фитопланктоном [Покровская и др., 1983]. В этих озерах усиление антропогенного эвтрофирования проявляется в сукцессии макрофитов. Свидетельством приближения критического уровня нагрузки является массовое развитие нитчатых водорослей. Дальнейшее возрастание эвтрофирующего воздействия приводит к срыву макрофитной экосистемы, резкой активизации фитопланктона и переходу экосистемы к обычному фитопланктонному типу. При нарастании эвтрофирующего воздействия происходят закономерные изменения в видовом составе макрофитов (главным образом, погруженных) и в степени зарастания водоемов [Olhe, 1958, Гигевич, 1991]. В олиготрофных и слабоминерализованных водоемах с прозрачной водой среди погруженных макрофитов господствуют низкорослые растения: мхи, ивоэтиды (полушниковые), при более высокой минерализации - харовые водоросли. При умеренном возрастании трофии, в мезотрофных озерах биомасса погруженной растительности и занимаемые ею площади максимальны, видовой состав их здесь наиболее богат, преобладают высокорослые элодеиды (элодея, роголистник, лютик, уруть, широколистные рдесты), вытесняющие низкорослые харовые водоросли (например, в озере Нарочь). Некоторые высокорослые виды харовых водорослей более устойчивы к повышению уровня трофии. В целом можно согласиться с мнением, что макрофиты мало пригодны для экспресс-анализа состояния водоемов, но могут быть очень полезны для целей мониторинга или при сравнении трофического статуса разных озер. [Доценко и др., 1991]. Степень покрытости водоема макрофитами. Связь этого показателя с процессами эвтрофирования несомненна, однако, его эффективность при оценке трофического состояния зависит от особенностей морфометрических характеристик водоема, прежде всего, площади мелководий. Помимо индикаторных организмов для оценки трофического состояния водоемов используются различные соотношения биологических показателей, например, отношение бактериопланктона к фитопланктону и др., которые отражают изменения в экосистеме водоема при ее эвтрофировании. В частности, известная реакция в изменениях зоопланктонного сообщества при повышение трофии позволяет считать хорошим индикатором соотношение мелких и крупных форм зоопланктона, а также соотношение рачкового планктона и коловраток в водоеме. В этом случае указанные соотношения отражают изменения пищевых связей в экосистеме при повышении ее трофии.
2. Гидрохимические показатели Поступление биогенных веществ в водоем. Величина нагрузки водоема биогенными веществами – один из главных параметров распространенных балансовых моделей эвтрофирования. Существует, однако, одна труднопреодолимая проблема при использовании этого показателя – необходимость разделения биогенной нагрузки на внешнюю (водосборную) и внутреннюю (водоемную). Это можно сделать только на основе специальных исследований роли внутриводоемных источников биогенных веществ в общем их круговороте в водоеме. Концентрация биогенных веществ. Содержание в воде водоемов биогенных веществ, в первую очередь общего содержания фосфора и азота, представляется очень эффективным показателем трофического состояния водоемов. Практически любое лимнологическое обследование водоема начинается с определения уровня содержания биогенных веществ в воде водоема. Классификации трофического состояния по содержанию общего фосфора представлены в табл. 1.4.
Таблица 1.4 Классификации типов водоемов по содержанию общего фосфора (мкг/л)
|