Морфология митотических хромосом
Как интерфазные, так митотические хромосомы состоят из элементарных хромосомных фибрилл — молекул ДНП (дезоксирибонуклеопротеида). На каждую хромосому приходится одна гигантская фибрилла ДНП, сложно уложенная в относительно короткое тельце — собственно митотическую хромосому. В митотической хромосоме существуют боковые петли этой гигантской молекулы дезоксирибонуклеопротеида. Боковые петли хромосом в вытянутом состоянии могут достигать 30 мкм. При их компактизации (спирализации) образуются структуры промежуточного характера — так называемые хромонемные фибриллы. Взаимодействие этих компонентов хромосом друг с другом и их взаимная агрегация приводят к конечной компактизации хроматина в виде митотической хромосомы.
Морфологию митотических хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе и в начале анафазы. Хромосомы в этом состоянии представляют собой палочковидные структуры разной длины с постоянной толщиной. У большинства хромосом удается легко найти зону первичной перетяжки (центромеры), которая делит хромосому на два плеча. Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины — субметацентрическими. Палочковидные хромосомы с очень коротким, почти незаметным вторым плечом называют акроцентрическими. В области первичной перетяжки расположен кинетохор. От этой зоны во время митоза отходят микротрубочки клеточного веретена, связанные с перемещением хромосом при делении клетки. Некоторые хромосомы имеют вторичные перетяжки, располагающиеся вблизи одного из концов хромосомы и отделяющие маленький участок — спутник хромосомы. Вторичные перетяжки называют ядрышковыми организаторами, т.к. именно на этих участках хромосом в интерфазе происходит образование ядрышка. В этих местах локализована ДНК, ответственная за синтез рибосомных РНК.
Плечи хромосом оканчиваются теломерами — конечными участками. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию. В каждом цикле деления теломеры клетки укорачивается, из-за неспособности ДНК-полимеразы синтезировать копию ДНК с самого конца. Данный феномен носит название концевой недорепликации и является одним из важнейших факторов биологического старения. Специальный фермент теломераза при помощи собственной РНК-матрицы достраивает теломерные повторы и удлиняет теломеры. В большинстве дифференцированных клеток теломераза заблокирована, однако активна в стволовых и половых клетках. Размеры хромосом, как и их число, у разных организмов варьируют в широких пределах. Совокупность числа, размеров и особенностей строения хромосом называется кариотипом данного вида.
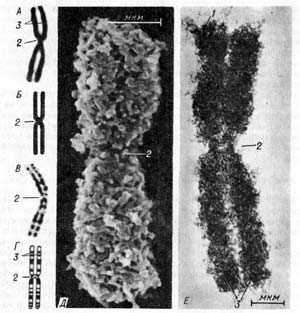
Строение хромосомы. Хромосома в световом микроскопе (А) и ее схематическое изображение (Б); хромосома при дифференциальной окраске (В) и ее схематическое изображение (Г); Д — хромосома в сканирующем электронном микроскопе; Е — хромосома в трансмиссионном мегавольтном электронном микроскопе; 1 — теломеры; 2 — центромеры; 3 — плечи хромосомы.
|