Синоатриальный(СА), синоаури-кулярный, или синусовый, узел
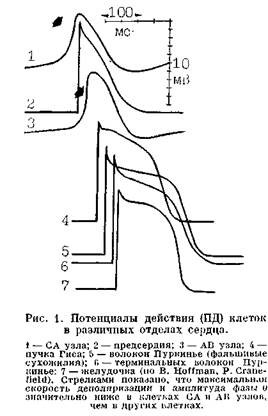
[Keith A., Flack M., 1907]. Узел находится в стенке правого предсердия, в большинстве случаев несколько лате-ральнее устья верхней полой вены [Синев А. Ф., Крымский Л. Д., 1985; Михайлов С. С., 1987; James Т., 1961; Anderson R. et al., 1981]. Его передняя часть («голова») лежит субэпи-кардиально у начала пограничной борозды (sulcus terminalis), задне-нижняя часть («тело» и «хвост») постепенно внедряется в мускулатуру пограничного гребня (crista terminalis) по направлению к нижней полой вене. Длина СА узла у взрослого человека — от 10 до 18 мм, ширина центральной части — 3—5 мм, толщина— 1—2мм. Хотя в кровоснабжении СА узла имеются индивидуальные различия, более чем у половины людей через его центр проходит одна сравнительно большая артерия, которая продолжается в стенке правого предсердия либо заканчивается в узле. Артерия СА узла является прямым продолжением первой пред-сердной ветви, отходящей у 60—70% людей от правой венечной артерии; у остальных людей артерия СА узла берет начало от огибающей ветви левой венечной артерии [Травин А. А. и др., 1982; Михайлов С. С., 1987]. Центральную часть СА узла называют «компактной зоной», в ней сосредоточены клетки длиной от 5 до 10 ммк, имеющие округлую или овальную форму. В световом и электронном микроскопах они кажутся бледными (Р-pale), поскольку в их цитоплазме мало миофибрилл, митохондрий и трубок саркоплазматиче-ского ретикулума. Эти узловые Р-клетки собраны в окруженные мембраной структуры, напоминающие гроздья винограда [James Т., 1961, 1965, 1985; Masson-Pevet M. et al., 1979, 1980]. Максимальный диастолический потенциал (потенциал покоя) клеток компактной зоны низкий: — 50 мВ, поэтому им свойствен медленный электрический ответ, т. е. низкая максимальная скорость деполяризации (Утах^й В/с), небольшая амплитуда и малая реверсия фазы 0 потенциала действия (ПД) (рис. 1). ПД практически не чувствителен к тетродотоксину (ТТХ) — веществу, блокирующему быстрые Na-каналы, которые, правда, имеются в клеточ ной мембране, но находятся здесь в инактивировэнном состоянии. По-видимому, единственным током, формирующим в этих клетках фазу О ПД, является медленный входящий ток isi, переносимый ионами Са++ и Na+. Ток isi угнетается блокаторами медленных каналов мембраны: ионами Mn++, Ni++, а также верапамилом. Восстановление инактивированного тока isi растягивается на несколько сотен миллисекунд. Соответственно этим особенностям ПД скорость проведения импульса в центральной части С А узла невелика: ее максимальная величина, измеряемая по длинной оси узловых клеток, составляет 0,05—0,2 м/с: их рефракторный период имеет большую продолжительность, чем ПД. «Доминирующее пейсмекерноо место» находится в центре компактной зоны, в участке площадью 0,3 см2. Именно здесь найдены группы узловых клеток (^5000) с синхронной электрической активностью и с паи-
большей скоростью спонтанной диа-стояической деполяризации, т. е. с самым высоким уровнем автоматизма. По направлению от центра к периферии СА узла сосредоточены удлиненные переходные Т-клетки (transitional), промежуточные по своим размерам и сложности между «узловыми» и сократительными предсердны-ми клетками. Они расположены более регулярно, в них возрастает число миофибрилл, больше имеется различных межклеточных соединений. Морфологические изменения переходных клеток коррелируют с их постепенной эволюцией от автоматического к неавтоматическому типу. Потенциал покоя этих клеток в абсолютном значении возрастает (от —60 до —65 мВ), увеличивается Vmax (в нижней части СА узла >10 В/с), повышаются амплитуда фазы 0 и ее чувствительность к ТТХ, понижается скорость (крутизна) спонтанной диастолической деполяризации [Blee-ker W. et al., 1980]. В СА узле много коллагена [Ор-thof Т. et al., 1987]. Вокруг узла имеется оболочка (СА-соединение), составленная из перинодальных волокон, которые по своим электрофизпо-логическим свойствам тоже занимают промежуточное положение между узловыми и сократительными клетками. Благодаря тому, что рефрактер-ность в узловых клетках сохраняется дольше, чем в предсердных (здесь она соответствует длительности ПД), преждевременные предсердные импульсы блокируются в перинодальных волокнах либо в самом СА узле. СА узел богато иннервирован хо-линергическими и адренергически-ми волокнами. На его автоматическую функцию воздействуют правосторонние блуждающий и симпатический нервы, вызывающие отрицательный и положительный хроно-тропные эффекты соответственно [Balsano F., Marigliano V., 1981; Ran-dall W., Ardell J., 1985]. Наибольшая чувствительность к ацетилхолину отмечается в центральной части СА узла, где, по-видимому, возрастает плотность мускариновых холиноре-цепторов [Bonke F. et al., 1984]. Ацо-тилхолин замедляет проводимость в центральной части СА узла и заметно удлиняет в ней период рефрактер-пости. Норадрепалин не изменяет скорости проведения импульса в СА узле, но укорачивает функциональный рефрактерпый период (ФРП) в центре СА узла и в перинодальпых волокнах. Медиаторы вегетативных нервов (наряду с пассивными электрическими свойствами клеток и межклеточными соединениями) оказывают существенное влияние на «пластичность» СА узла, его способность реагировать на самые различные раздражители [Bukauskas F. ol al., 1987; Bonke F. et al., 1985]. Здесь следует коснуться вопроса о взаимодействии блуждающего и симпатического нервов в регулировании выработки синусового импульса и его распространения от центра к перифо- рии узла и к миокарду предсердий. Окончания вегетативных нервов расположены очень близко друг к другу, что способствует парасимпатическому-симпатическому взаимодействию как на пресипаптическом, так и па постсинаптическом уровне. Лцетил-холип выделяется из окончаний блуждающего нерва порциями в определенной фазе сердечного цикла. При изменениях длины цикла и некоторых других обстоятельствах аце-тилхолштовый «залп» может смещаться во времени. В одних случаях (на фоновом усилении симпатической активности) это приводит к более сильному торможению выделения норадреналипа и его положительного хропотроппого эффекта, что получило название акцентуированного антагонизма с общим усилением ингибиторного вагуспого эффекта [Levy M., Martin P., 1984]. В других случаях ацетилхолип может усиливать положительный хротютропный эффект симпатических стимулов в СА узле [Yang Т., LevyM., 1984; Wal-lickD. etal., 1986]. - Миокард предсердий. Он имеет сложное строение, и клеточная структура правого предсердия не совпадает со структурой левого предсердия. Эти цитологические, а также функциональные (электрофизиологические) особенности мышцы правого предсердия, вероятно, связаны с тем, что именно оно прилежит к АВ узлу, и переход импульсов от СА узла к АВ узлу осуществляется по правому предсердию. Здесь мы коснулись все еще не регаеттпого вопроса о путях ускоренного проведения в правом предсердии. Напомним, что выдвинуты три основные концепции движения синусовых импульсов в правом предсердии. Первая из них, сформулированная еще Th. Lewis и соавт. (1910), основывается на признании равномерного радиального распространения волны возбуждения в предсердной мышце. Иной точки зрения придерживаются R. Anderson и соавт. (1981), а также некоторые другие исследователи. Они полагают, что в определенных участках правого предсердия обеспечивается «предпочтительное проведение импульсов» (этот термин значительно раньше применил J. Eyster, 1914, 1916), т.е. более быстрое их проведение, благодаря особому расположению (геометрии) обычных сократительных мышечных пучков. Третью концепцию о существовании в предсердиях специализированных быстрых путей (трактов) проведения многие годы развивают Т. James и соавт. (1963—1985). Согласно их представлениям, короткий передний [Bachmann G., 1916] и средний [Wenckebach К., 1907, 1908] тракты, покинув СА узел, опускаются вниз по межпредсердной перегородке и присоединяются к верхнему краю АВ узла. Более длинный задний тракт [Thorel С., 1909; James Т., 1963] начинается от заднего края СА узла, проходит вдоль пограничного гребня к клапану Евстахия и затем в межпредсердной перегородке, над коронарным синусом, соединяется с нижнезадней частью АВ узла. Почти одновременное возбуждение правого и левого предсердий обеспечивается ускоренным движением импульса от СА узла к левому предсердию по большому мышечному пучку Бахма-на, который рассматривается как ветвь переднего межузлового тракта. L. Sherf, T. James (1979) нашли в зоне межузловых трактов при электронной микроскопии 6 типов клеток: 1) Пуркинье-подобные клетки; 2) широкие переходные клетки; 3) тонкие переходные клетки: 4) Р-клетки; 5) амебоидные клетки; Пуркинье-подобные клетки («бедные мпофибрилами клетки»). Наконец, различные переходные клетки служат ср'еднескоростными проводниками импульса между очень медленно проводящими Р-клетками и быстро проводящими Пуркинье-подобпыми клетками. Взгляды Т. James разделяют М. Vassalle и В. Hoffman (1965), М. Lev и соавт. (1977), G. Pastelin и соавт. (1978), A. Waldo и соавт. (1979), S. Bharati и соавт. (1983) и другие исследователи. М. Hiraoka и II. Adaniya (1983) показали, в част-пости, что в правом предсердии кролика импульсы от GA узла к АВ узлу движутся быстро вдоль левой и правой ветви пограничного гребня, что соответствует предполагаемому положению переднего и заднего межузловых трактов. Перерезка этих ветвей сопровождается отчетливым замедлением проведения (на 33 мс), которое осуществляется уже соседними участками сократительного миокарда. При гистологическом исследовании в ветвях пограничного гребня были найдены Пуркинье-подобные клетки. W. Tse (1986) выявил в сердце собаки специализированный межузловой тракт, распространяющийся по свободной стенке правого предсердия и затем вдоль основания сеп-тальной створки трехстворчатого клапана. По мнению автора, значение этого тракта, составленного из переходных и паранодальных волокон, велико, поскольку при повреждении паранодальных волокон возникает АВ блокада. Еще раньше Н. Tsuchinashi и соавт. (1980) обнаружили в сердце кролика особые пути, связывающие нижний отдел левого предсердия с правым предсердием и АВ узлом. этрт специализированные волокна распределяются вдоль передне- п заднемедиального сегментов кольца митрального клапана. В норме импульсы отсюда, по-видимому, не поступают к желудочкам, так как их опережает волна возбуждения из правого предсердия. В условиях па- тологии эти нижние левопредсердныс пучки (если они существуют в сердце человека) могут быть источником аритмий. Представляют интерес последние работы, выполненные в этой области отечественными морфологами. В лаборатории С. С. Михайлова (1987) удавалось выявлять межузловые пути, правда, не в каждом исследованном сердце человека. Автор делает вывод, что в предсердиях существует более сложная система проводящих пучков, чем это представляется на основании данных литературы. А. Ф. Сипев, Л. Д. Крымский (1985) пришли к заключению, что «тракты» в виде мышечных пучков действительно можно обнаружить и выделить. Однако их микроскопическое строение не соответствует трем обязательным морфологическим критериям проводящей системы сердца: а) постоянству и структурной непрерывности «трактов» на всем протяжении от СА узла к атриовентрику-лярному (АВ) узлу; б) специфичности, т. е. их построению из специализированных кардиомиоцитов (истип-пых клеток Пуркинье); в) изоляции соединительнотканным футляром от окружающего сократительного миокарда желудочков. С дискуссией о характере проведения синусовых импульсов в предсердиях тесно связан вопрос о входах этих импульсов в АВ узел и соответственно о выходах из него. В 1978г. М. Janse и соавт., а затем Н. lirmma, L. Dreifus (1986) показали, что существуют два таких входа: 1) передний — из основания межпредсердной перегородки; 2) задний — со стороны пограничного гребня. Если синусовые импульсы обычно используют оба входа, равномерно проникая в АВ узел, то экстрасистолы (преждевременные предсердные экстрастимулы) и волны фибрилляции предсердий (ФП) попадают в АВ узел главным образом через тот вход, к которому они ближе [Mazgalew T. et al., 1984]. Такое активное проведение в АВ узел дополняется пассивным перинодаль- ным проведением через другой вход. От места и времени поступления преждевременного экстрастимула (экстрасистолы и т. д.) в АВ узел зависят рефрактерность, скорость и последовательность возбуждения его структур, появление неравномерной активации не только самого АВ узла, но и ствола пучка Гиса, а также развитие1 блокад. Заканчивая обсуждение вопроса о внутрипредсердных специализированных путях проведения, мы считаем нужным подчеркнуть, что большинство исследователей, признающих их существование, не рассматривают эти «тракты» в анатомическом смысле как непрерывные, отграниченные оболочкой структуры. В функциональном же отношении, т. е. по своим электрофизиологическим свойствам, составляющие тракты специализированные кардиомиоциты отличаются от окружающих их сократительных клеток способностью более быстро проводить импульсы, а также устойчивостью к гиперкалие-мип с сохранением синовентрикуляр-иого проведения, наконец, своеобразием реакции па адреналин [Магу-Rabine L. et al., 1978]. Разумеется, предсердпая система проведения не может приравниваться к впутриже-лудочковой проводящей системе Гиса — Пуркинье. Что касается сократительных пред-сердных клеток, то им свойствен быстрый электрический ответ: потенциал покоя равен —80 мВ, Vmax колеблется от 80 до 300 В/с, фаза О ПД имеет выраженную крутизну, большую амплитуду и реверсию. Эта фаза генерируется быстрым входящим Na-током (1ыа), который можно пода-пить ТТХ. Скорость проведения импульса в сократительных предсерд-пых волокнах — 0,6—0,7 м/с, тогда как расчетная скорость движения по «трактам» составляет i— 1,5 м/с. В сократительном миокарде предсердий рассеяны очаги латентного автоматизма. W. Randall и соавт. (1978) обнаружили у 80% животных; после разрушения GA узла спонтан- ную электрическую активность в области евстахиева клапана. Вслед за введением изопропилнорадреналипа у большинства из этих животных водитель ритма смещался к межпред-сердному пучку Бахмана. В хронических опытах на собаках та же группа исследователей установила, что предсердным центрам автоматизма свойственна нерегулярная активность, которая становится более правильной под воздействием атропина сульфата. Учащение эктопического ритма в ответ па физическую нагрузку удавалось предотвратить с помощью блокатора 8-адрепергических рецепторов. Стимуляция симпатических нервов смещала водитель ритма к нижней части пограничного гребня, где предположительно проходит задний межузловой «тракт». L. Mary-Rabine и соавт. (1978) регистрировали в ткани правого предсердия, резецированной у больных во время операции на сердце, ПД, образование которых замедлялось при добавлении адреналина в низкой концентрации и ускорялось при более высоких концентрациях адреналина. Эффект низких концентраций адреналина блокировался а-адреноблока-тором фептоламином; эффект высоких концентраций — р-адреноблока-тором пропранололом, что сходно с реакциями, происходящими в желудочковых волокнах Пуркипьо. Очевидно, что в мышце предсердий имеются а- и р-адренорецепторы. В 1981 г. J. Loeb, J. Moran показали, что у собак при перфузии артерии СА узла ацетилхолииом или фи-зостигмипом (ингибитор холинэсте-разы) происходило демаскироваппе скрытых водителей ритма, которые оказались более чувствительными к парасимпатическим влияниям, чем СА узел. Последняя реакция может быть дополнительным механизмом, обеспечивающим главенствующее положение СА узла в предсердиях. Уровень автоматизма скрытых предсерд-пых водителей ритма достигал 73% синусового автоматизма. Наиболее раннюю активацию предсердий npir торможении СЛ узла авторы отметили вблизи заднего межузлового «тракта». Группа авторов во главе с J. Воь ncau (1978, 1985) придерживается мнения, что множественные пред-сердпые водители ритма не являются полностью изолированными, а образуют вместе с СА узлом систему, названную ими «предсердным пейсме-керным комплексом». В правом предсердии у собаки этот комплекс включает очаги автоматизма, которые распространяются в каудокраниальном направлении на протяжении 40— 50 мм, вблизи от пограничной борозды, между верхней и нижней полыми венами. Ширина зоны пейсмекеров составляет около 15 мм, а площадь — 45X15 (675 мм2), что в 10 раз больше размеров СА узла. Самые быстрые, краниальные, водители ритма располагаются позади rostrum СА узла в области Remak [Remak R., 1844], содержащей большое количество гап-глиозных клеток. Самые медленные, каудальные, водители ритма сосредоточены в зоне, известной как область Fredericq [Fredericq L., 1906]; она соответствует месту, где находятся предсердные водители ритма, продолжающие возбуждать умирающее сердце (ultium moriens). Особенность предсердного пейсмекерного комплекса состоит в том, что возбуждение может одновременно (или лишь с минимальным запаздыванием) начинаться по меньшей мере в двух центрах автоматизма. Благодаря такому взаимодействию обеспечивается быстрая деполяризация большого участка миокарда предсердий, например 900 мм2 за первые 10 мс. Как отдельные водители ритма, так и весь пейсмекерный комплекс регулируются парасимпатическими и симпатическими нервами [Boineau J. et al., 1985]. Недавно J. Boineau и соавт. (1988) представили данные о существовании предсердного пейсмекерного комплекса в сердце человека. Автоматическая и триггерная активность выявляется в коронарном синусе, имеющем отношение к элек- трогемозу левого предсердия [Wil A., Cranei'ield P., 1977]. Кроме того, в левом предсердии обнаружены автоматические клетки вокруг устьев четырех легочных вен и в пучке Бахмапа. В 1979 г. A. Wit и соавт. установили, что у человека и у экспериментальных животных мышечные клетки двустворчатого клапана обладают способностью спонтанно вырабатывать импульсы. По своей ультраструктуре эти клетки не отличаются от обычных предсердных клеток, однако им свойственна диастолическая деполяризация. Генерация импульсов опосредуется медленными каналами мембраны, поскольку она подавляется верапамилом и нечувствительна к ТТХ. При электрической стимуляции клеток и добавлении к ним адреналина либо при воздействии сердечных гликозидов появляются задержанные постдеполяризации и триггерная активность, устраняемая ацетилхолином и верапамилом. Таким образом, в створках митрального клапана могут при благоприятных условиях формироваться как медленные автоматические, так и более быстрые триггерные ритмы [Ульянинский Л. С., Кашарская И. Л., 1981; Wit A. et al., 1979]. G. Rozanski и J. Jalife (1986) обнаружили, что такими же свойствами обладают мышечные клетки трехстворчатого клапана (в сердце кролика). Как митральный, так и трехстворчатый клапаны богато иннервированы парасимпатическими и симпатическими нервными волокнами. Адренергичес-кие и приходящие в определенное время холинергические стимулы способны увеличивать наклон диастоли-ческой деполяризации в клетках створчатых клапанов (парасимпатическое-симпатическое взаимодействие), т. е. усиливать их автоматизм. Атриовентрикулярный узел Ашоф-фа —Тавара [Tawara S., 1906; Aschoff L., 1908, 1910]. Он расположен в задней части межпредсердной перегородки, справа под эндокардом, впереди от устья коронарного синуса, непосредственно выше места при-
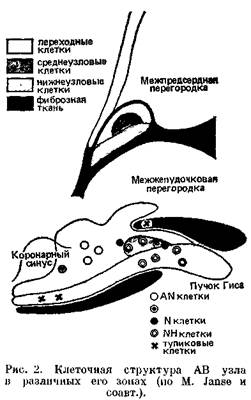
крепления к перегородке септальнои створки трехстворчатого клапана; иначе говоря, — на правом треугольнике центрального фиброзного тела сердца [Синев А. Ф., Крымский Л. Д., 1985; Михайлов С. С., 1987; Andersen R. et al., 1981; Bharati S. et al., 1983]. Оперирующие на проводящей системе сердца хирурги часто определяют положение АВ узла по треугольнику Коха [Koch W., 1912], в переднем углу которого находится АВ узел. Передневерхней стенкой треугольника Коха служит сухожилие Тодара (фиброзный пучок, проходящий в основании клапана нижней полой вены); нижнюю стенку образует место прикрепления к фиброзному кольцу септальнои створки трехстворчатого клапана; задневерх-нюю стенку составляет устье коронарного синуса [Anderson R. et al., 1980]. При гистологическом и гистохими-ческом исследованиях выявляется трехслойное строение АВ уела, составленного из клеток, различающихся формой, размерами, контактами и, наконец, скоростью проведения импульса (рис. 2). Некоторые исследователи сравнивают структуру АВ узла с лабиринтом из специализированных волокон [Watanabe Y., Drei-fus L., 1965, 1980; Mazgalev Т. et al., 1986]. Проксимальный слой АВ узла, связанный с мышцей правого предсердия, представлен переходными (Т) клетками, удлиненными, по меньшими по величине, чем пред-сердные сократительные клетки. Группы переходных клеток отделены друг от друга прослойками кол-лагеновых волокон, резко замедляющих движение импульса. Н. Hecht и соавт. (1973) назвали этот слой преддверием АВ узла. Второй слой — собственно АВ узел, или компактный АВ узел ("Knotenpunkten" no S. Та-wara), имеет длину около 6 мм, ширину — 2—3 мм, толщину — 1,5 мм. Он составлен из плотно соприкасающихся клеток, среди которых, помимо переходных (Т) клеток, имеются округлые Р-клетки с малым числом органелл и миофибрилл. Их здесь заметно меньше, чем в GA узле [James Т., 1985]. Наконец, третий слой АВ узла — длинная дисталъная часть, непосредственно переходящая в атриовентри-кулярный пучок Гиса. В свое время A. Paes de Carvalho (1959) выделил три электрофизиологические зоны АВ узла (в сердце кролика), отличавшиеся особенностями потенциалов действия: AN (atrium-nodus), N (nodus), NH (nodus-His). Теперь известно, что ПД типа AN вырабатываются в слое переходных клеток; ПД типа N — в небольшой группе клеток среднего, компактного, слоя и частично в переходном слое АВ узла; ПД типа NH — в нижнеузловом слое; в этом же слое найдены группы клеток, образующие так называемые электрофизиологические тупики, в которых угасает проведение импульсов (см. рис. 2).
Сложность строения АВ узла и других образований, связывающих предсердия с желудочками, многообразие их функций явились причиной появления нового термина— «АВ соединение». Правда, между морфологами и клиницистами существуют расхождения в определении этого понятия. Первые его толкуют расширительно, включая: 1) переходный слой и паранодальные волокна; 2) компактный АВ узел и его дис-тальную часть; 3) проникающий АВ пучок и 4) ветвящийся АВ пучок. Надо признать, что основания для такой трактовки дают некоторые эмбриологические данные. Однако, с клинической (электрокардиографической) точки зрения, к АВ соединению следует относить только первые три образования — до места разделения общего ствола на ветви. Соответственно проникающий АВ пучок входит в наджелудочковую область, а ветвящийся АВ пучок — в желудочковую. В последующем мы применяем термин «АВ соединение» именно в таком смысле. У 80% мужчин и 93% женщин АВ узел снабжается кровью из ветви правой венечной артерии; у остальных людей кровь к АВ узлу поступает из огибающей ветви левой венечной артерии [Травин А. А. и др., 1982; James Т., 1961]. Важную коллатераль к АВ узлу образует артерия Кугеля [Kugel М., 1927]. АВ узел богато ин-нервирован парасимпатическими и симпатическими волокнами. Как и в СА узле, здесь имеется односторонняя избирательность иннервации: только левый блуждающий нерв вызывает отрицательный дромотропныи эффект в АВ узле; только левый симпатический перв укорачивает время АВ узлового проведения без воздействия на длину интервала Р—Р [Bal-sano F., 1981; Marigliano V., 1981; Randall W., Ardell J., 1985]. Наибольшая плотность нервных окончаний и рецепторов (в частности, мускарино-вых) отмечается в зоне N и в прилегающей к ней части зоны NH АВ узла [Mazgalew Т. et al., 1986]. На про- ведение импульса через АВ узел оказывают влияние вариации парасимпатического-симпатического взаимодействия, упоминавшегося выше [Ur-thaler F. et al., 1986]. В физиологических условиях существует параллельный регуляторный контроль частоты синусового ритма и АВ узловой проводимости, направленный на поддержание АВ проведения 1:1 [Ви-trous G. et al., 1987]. Однажды Н. Marriott (1966) назвал АВ узел «островом чудес в океане неизвестного». Это все еще актуальное определение не только отражает неполноту наших сведений об АВ узле, но и содержит признание важной роли, которую играет АВ узел в системе проведения и образования импульса. Основные функции АВ узла: 1) физиологическая задержка импульсов, движущихся от предсердий к желудочкам (синхронизация сокращений предсердий и желудочков с задержкой); 2) фильтрация (сортировка) предсердных волн возбуждения, препятствующая слишком частой активации желудочков; 3) защита желудочков от раннего, преждевременного возбуждения в уязвимой фазе их цикла. О степени физиологической анте-роградной АВ задержки можно судить по следующим показателям: на преодоление импульсом небольшого АВ узла требуется от 50 до 130 мс (интервал А — Н Гис-электрограм-мы), т. е. намного больше времени, чем на прохождение длинного пути в правом предсердии. Еще медленнее осуществляется ретроградное вентрикуло-атриальное (ВА) узловое проведение (интервалы Н — А и V — А Гис-электрограммы). Например, у свиньи при стимуляции правого желудочка время ВА узлового проведения в среднем составляет 178±8,5 мс против 120± 18 для анте-роградного АВ узлового проведения [Bowman Т., Hughes N., 1984]. У 20% людей с неизмененным антероград-ным АВ проведением отсутствует ВА узловое ретроградное проведение [Mahmud R. et al., 1985]. Правда, во многих случаях ото связано с усиленным воздействием блуждающего нерва: после введения атропина ВА проведение восстанавливается. При ретроградном движении импульса через АВ узел прежде всего возбуждается нижнеперегородочная часть правого предсердия (иногда — область устья коронарного синуса — Gallagher J., 1987), затем все правое предсердие и с минимальным запаздыванием — левое предсердие. Крайне низкая скорость распространения импульса в АВ узле (в среднем 0,05 м/с) зависит от преобладания в нем клеток с медленным электрическим ответом. В зоне N максимальный диастолический потенциал этих клеток равен — 60 мВ, небольшая пологая фаза О ПД устойчива к ТТХ, но блокируется ионами Мп++ и верапамилом. Между тем вопрос о месте наиболее выраженной задержки импульса в АВ узле все еще не вполне выяснен. Важным, хотя и не единственным, показателем скорости проведения импульса клетками является величина Vmax (dV/dt). По данным W. Tse (1986), Vmax в пара-подальных волокнах, подходящих к компактному АВ узлу, равняется 2,5 В/с, в самом АВ узле — 7 В/с, в стволе пучка Гиса — 30 В/с. Автор приходит к выводу, что АВ задержка волны возбуждения происходит в па-ранодальных волокнах у входа в АВ узел. Еще раньше М. Janse и соавт. (1976) указали, что от 20 до 80% времени А—Н и от 18 до 80% времени Н—А (Гис-электрограмма) приходится на прохождение зоны AN узла, что близко к данным W. Tse. Только 20% времени АВ проведения затрачивается на прохождение зоны N узла. Однако именно в этой небольшой зоне под влиянием усиливающейся вагусной стимуляции фрагментируется и утрачивает синхронность фронт возбуждения, что приводит к АВ узловым блокадам и, в частности, к развитию периодики Самойлова — Венкебаха. Характеризуя АВ узел, следует коснуться еще двух его функций. I Горная ил них имеет отношении к возможности продольного" разделения АВ узла на два электрофизиологических канала — основу для re-entry и кругового движения импульса (рассмотрению этого механизма мы уделили место ниже — в разделах о реципрокных комплексах и АВ узловых реципрокных тахикардиях). Вторая функция имеет отношение к автоматизму АВ узла. В течение последних 30 лет допускалось, что автоматическая активность свойственна только клеткам NH-зопы АВ узла. Основанием для такого заключения послужили микроэлектродные исследования В. Hoffman и P. Granefied (1960), в которых не удалось in vitro обнаружить спонтанную диастоличе-скую деполяризацию в клетках AN-и N-зоны АВ узла (в сердце кролика). Однако позже некоторые авторы смогли зарегистрировать диастоли-ческую деполяризацию в небольших изолированных препаратах АВ узла из сердца кролика [Kokuburn S. et al., 1980] и собаки [Tse W., 1986]. Недавно И. Ирисава и соавт. (1988) подтвердили в эксперименте, что клеткам АВ узла кролика присуща латентная пейсмекерная активность, однако в интактном сердце эти клетки никогда не возбуждаются спонтанно. A. Wit и P. Cranefield (1982) так объясняют эти противоречивые данные. В здоровом (интактном) сердце кролика действительно отсутствует автоматическая активность в зонах AN и N АВ узла. Если же связь предсердий с АВ узлом нарушается, то в этих участках АВ узла можно уловить образование автоматических импульсов, частота которых превышает частоту синусового ритма, что исключает механизм сверхчастого подавления центров АВ узла. Причины угнетения автоматизма АВ узла представляются иными: поскольку предсердные клетки имеют более отрицательный потенциал покоя, чем клетки АВ узла, то возникающий между ними ток приобретает такое направление, которое предотвраща- ст спонтанную диастолическую деполяризацию узловых клеток. Любой фактор, который ослабляет это элек-тротаническое межклеточное взаимодействие, будет способствовать проявлению скрытых автоматических потенций Л В узла (выраженный фиброз в место соединения правого предсердия и АВ узла и др. ' В 1983 г. R. Hariman и Chia-Maou С lien иришли к выводу, что у собак замещающий ритм АВ соединения может исходить из автоматических (Р) клеток зоны N АВ узла. Авторы напоминают о работе В. Scherlag и соавт. (1971), показавших, что оуаба-цн оказывает различное влияние на ритмы, берущие начало в АВ узле и стволе пучка Гиса. Пока не вполне ясно, в какой степени эти экспериментальные данные приложимы к патологии человека, но изучение замещающих ритмов у больных с врожденными полными АВ блокадами подтверждает возможность возникновения проксимальных и дистальных АВ ритмов. АВ пучок Гиса. Он служит продолжением дистальной части АВ узла [His W., 1893]. Начальный, или проникающий, сегмент пучка Гиса (общий ствол) длиной 10 мм проходит через центральное фиброзное тело в непосредственной близости от отверстий митрального и трехстворчатого клапанов и направляется вперед по верхнему краю мембранозной части мсжжолудочковой перегородки. Кол-лагеповые прослойки разделяют общий ствол на множество продольных пучков, слабо связанных друг с другом в поперечном направлении. Это обеспечивает разделение широкого фронта возбуждения на большое число равномерно движущихся изолированных волн, каждая из которых, по-видимому, достигает определенного предназначенного для нее участка миокарда желудков. Продольная диссоциация пучка Гиса, т. е. нарушение синхронности движения импульса, может имитировать изменения внутрижелудочковой проводимости [NarulaO., 1977, 1987]. Начинающийся у нижнего края мембранозной части межжелудочковой перегородки ветвящийся сегмент пучка Гиса представлен двумя ножками: правой и левой. Правая ножка образует изолированную ветвь длиной 4,5—5 см, направляющуюся вперед и вниз к различным субэндокар-диальным участкам правого желудочка и межжелудочковой перегородки. Первый функциональный контакт веточки правой ножки с сократительными волокнами происходит у основания передней сосочковой мышцы правого желудочка. Левая ножка чаще начинается с широкого основания, лежащего суб-эндокардиально на левой стороне мышечной части межжелудочковой не-регородки. Направляясь слегка вперед и вниз, она вскоре разделяется на три главных разветвления, на что обратил внимание еще S. Tawara (рис. 3). В настоящее время большинство исследователей придерживаются мнения, что левая ножка (рис. 4) имеет мыоговар'иантное, трех-нучковое строение [Massing G. и James Т., 1976; Kulbertus Н. и Demou-lin J., 1977]. Тонкое и протяженное передневерхнее разветвление левой ножки (длина — 2—2,5 см, толщина — 3 мм) подходит к корню и середине передней сосочковой мышцы. Широкое и более короткое заднениж-нее разветвление (толщина — 6 мм), которое как бы служит продолжением общего основания ножки, распространяется к началу задней сосочковой мышцы. Центральное, или собственно переднее, разветвление левой ножки чаще берет свое начало из угла, образованного двумя другими разветвлениями, либо от одного из них. Оно направляется вниз к средней части межжолудочковой перегородки, поэтому его нередко называют среднеперегородочным (третьим) разветвлением левой ножки. Между ветвями левой ножки имеется широкая сеть анастомозов как в проксимальных, так и в дистальных участках [Lev M. et al., 1977]. Основываясь на гистопатологических данных, по-
Рис. о. Фотография <• оригинального рисунка S. Tawara — распространение ветвей левой ножки в сердце человека [Tawara S. Des Reizlei-lungsystem der Saugo-tierherzens, 1906].
10,JV,.Ж ir-^w ir/a
18' Рис. 4. Схема распространения волокон (ветвей) левой ножки в 20 сердцах людей, не страдавших заболеваниями миокарда (по J. Demoulin, H. Kulbertus). лученных Y. Nakaya и соавт. (1987), D. Spodick (1988) называет проводящую систему левого желудочка многопучковой: «подобно вееру она широко распространяется по левой стороне межжелудочковой перегородки». При этом «двухпучковая концепция» М. Rosenbaum и соавт. (1970) сохраняет свое клинико-элек-трокардиографическое значение (см. гл. 16). Разделяясь на все более мелкие веточки, ножки пучка Гиса заканчиваются сетью волокон Пуркинъе [Pur-kinje J., 1845], которая является последним звеном специализированной проводящей системы сердца. Клетки Пуркинье проникают во внутренние 2/з мышечной стенки желудочков; субэпикардиальный слой почти лишен их [Kaplinsky E. et al., 1980]. Соединение клеток Пуркинье с сократительными клетками происходит без синапсов через переходные (Т) клетки [Tan R. et al., 1989]. С электрофизиологических позиций вполне оправданно объединение пучка Гиса и его ножек с их конечными разветвлениями в систему Гиса — Пуркинъе. Составляющие эту систему клетки отличаются быстрым
электрическим ответом [Баркинблит М. Б. и др., 1982; Розенштраух Л. В., 1982] и имеют следующие характеристики: максимальный диастоличе-ский потенциал (потенциал покоя) равен —88 ±1,7 мВ, Vmax колеблется от 30 до 200—600 В/с, амплитуда фазы О ПД достигает 118±1,9 мВ, длительность ПД равна 376 ±10,8 мс; клеткам свойственна спонтанная диастолическая деполяризация [Der-sham G., Han J., 1980]. Скорость проведения импульса нарастает от 1,5 м/с (в общем стволе) до 3—4 м/с (в клетках Пуркинье). В физиологических условиях в системе Гиса — Пуркинье нет декрементного (затухающего) проведения. На электрограмме пучка Гиса (ЭПГ) возбуждение общего ствола отражается потенциалом Н продолжительностью 15— 20 мс. Интервал Н—V Гис-электро-граммы соответствует времени движения импульса по ножкам пучка Гиса и сети Пуркинье до начала возбуждения миокарда желудочков; в норме он равен 30—55 мс. В проводящей системе желудочков имеется привратниковый (шлюзо-лый) механизм, функция которого определяется различиями в продолжительности ПД клеток Пуркинье. Зона наибольшей продолжительности ПД и, следовательно, рефрактер-ности расположена на 2—3 мм прок-сим
|