Головна сторінка Випадкова сторінка
КАТЕГОРІЇ:
АвтомобіліБіологіяБудівництвоВідпочинок і туризмГеографіяДім і садЕкологіяЕкономікаЕлектронікаІноземні мовиІнформатикаІншеІсторіяКультураЛітератураМатематикаМедицинаМеталлургіяМеханікаОсвітаОхорона праціПедагогікаПолітикаПравоПсихологіяРелігіяСоціологіяСпортФізикаФілософіяФінансиХімія
Соуси без борошна
Дата добавления: 2015-10-12; просмотров: 1336
|
|
Одноклеточные организмы имеют упругую оболочку, которая за счет тургора обеспечивает постоянную форму и играет роль «наружного скелета». Живые клетки многоклеточного организма сохранили эту особенность. У водных многоклеточных растений, обладающих тонкими клеточными оболочками, роль фактора, поддерживающего форму организма и обеспечивающего его прочность, играет вода. Для наземных организмов, многие из которых обладают крупными размерами, такая опорная система оказалась малоэффективной; у них возникли специализированные механические ткани, состоящие из клеток с утолщенными оболочками, которые выполняют опорную функцию даже при отмирании протопласта.
Функцию «внешнего скелета* у крупных древесных растений до некоторой степени выполняют перидерма и корка, мощный слой опробко-вевших тканей, одевающих растение снаружи. Но только механические ткани, появившиеся у растений в результате приспособления к наземной жизни, взаимодействуя с другими тканями организма, образуют прочный «внутренний скелет* растения, его арматуру. Поэтому механические ткани часто называют арматурными. В некоторых учебных пособиях систему механических тканей называют стереомом (греч. stereos — твердый, прочный), а составляющие ее клетки — етереи-дами.
Степень развития механических тканей во многом зависит от условий обитания: они почти отсутствуют у растений влажных лесов, у многих прибрежных растений, но зато хорошо развиты у большинства растений засушливых местообитаний — склерофитов.
Все механические ткани по ряду признаков можно разделить на несколько групп, каждая из которых специфична по строению. Таких основных групп три: колленхима, склеренхима и скле-реиды.
КОЛЛЕНХИМА
Колленхима (греч. кона — клей и ёпспуша, букв.: налитое, здесь — ткань) — паренхимная ткань, на поперечном разрезе клетки ее имеют разнообразную форму, близкую к 4—5-гранной, на продольном — несколько вытянуты по оси (рис. 88, А, Б). Колденхима появляется только как первичная ткань и служит для укрепления
|
|
|
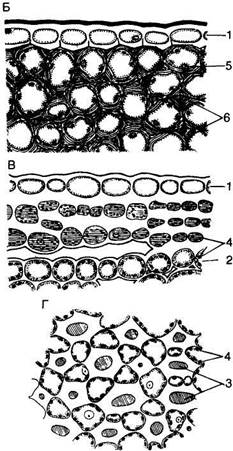
Рис. 88. Колленхима:
А — продольный разрез уголковой колленхимы листового черешка шалфея (Salvia sclarea): a — в световом, б — в электронном микроскопе (схемах); 1 — простые поры; 2 — плазмодесмы; 3 — центральная вакуоль; 4 — митохондрии; 5 — хлоропласта; 6 — срединная пластинка; Б—Г — основные типы колленхимы на поперечном срезе: Б — уголковая колленхима черешка листа плюща (Hoya camosa); В — пластинчатая колленхима черешка астранции (Astrantia major); Г — рыхлая колленхима черешка листа белокопытника (Petasites niveus); I — эпидермис; 2 — хлорофиллоносная паренхима; 3 — межклетники; 4 — хлоропласта; 5 — ядро; 6 — утолщения оболочки клеток
молодых стеблей и листьев, когда еще продолжается растяжение клеток в длину. Обеспечивая щючиость органу, колленхима в то же время слособна растягиваться по мере роста молодых органов. Пластичное (остаточное) растяжение оболочек возможно лишь при активном воздействии живого содержимого, которое выделяют вещества, снижающие упругость оболочек. Такой способностью могут обладать только живые клетки. Как показывают новейшие исследования, оболочка клеток колленхимы, толщина которой может достигать 5—6 мкм, характеризуется высоким содержанием целлюлозы (около 30 %) и ге-мицеллюлоз (свыше 50 %), воды, бедна пектиновыми веществами, лигнин в ней обычно отсутствует. Особенность оболочки — многослойность, видимая под электронным микроскопом как многочисленные концентрические слои (ламеллы) с резким изменением ориентации откладываемых целлюлозных микрофибрилл (продольных и поперечных). Содержание микрофибрилл в каждом слое более или менее одинаковое, они погружены в матрикс из целлюлоз.
Особенность живых клеток колленхимы, часто содержащих даже хлоропласты, состоит в том, что выполнять свое назначение арматурной ткани она может только в состоянии тургора. При потере молодыми органами воды тонкие участки оболочек складываются гармошкой, листья и стебли теряют упругость и обвисают, т. е. завядают.
Располагается колленхима почти всегда по периферии органа или сплошным слоем, или собрана отдельными пучками, связанными с проводящей тканью. В стеблях она часто в виде компактных пучков тянется вдоль стебля, например у ребристых стеблей зонтичных.
Колленхима относится к простым тканям, поэтому границы ее обычно очерчены довольно резко. Однако иногда образуются переходные формы от колленхимных клеток к типичной паренхиме.
Клетки колленхимы характеризуются неравномерным утолщением стенок (слоистость, о которой говорилось выше), за счет чего они могут выполнять опорную функцию. На рис. 88, А, б показано электронно-микроскопическое строение типичной колленхимной клетки. Хорошо видна слоистая утолщенная оболочка, в которой встречаются редкие простые поры (рис. 88, А, б, I), пронизанные многочисленными плазмодесмами (рис. 88, А, б, 2). Клетки колленхимы рано вакуо-лизируются: уже в самом начале дифференциации (отложение утолщений) образуется центральная вакуоль (рис. 88, А, б, 3), однако слой постенной цитоплазмы, куда перемещается и ядро, до прекращения роста остается шире, чем в соседней основной ткани. Ядро обычно крупное, а цитоплазма насыщена органоидами, главным образом митохондриями и диктиосомами.
Характерное утолщение стенок клеток колленхимы носит специальное название — утолщение колленхиматозного типа. В зависимости от характера утолщений стенок и соединения клеток между собой различают уголковую, пластинчатую и рыхлую колленхиму.
В уголковой колленхиме, наиболее распространенной, оболочка сильно утолщается в углах, где сходятся соседние три-пять клеток. Утолщенные части оболочек обычно сливаются между собой, так что границы отдельных клеток обнаруживаются с трудом (см. рис. 88, А, Б).
В пластинчатой колленхиме сплошными параллельными слоями утолщаются тангенциальные стенки, радиальные же остаются тонкими. Чаще всего эти слои параллельны поверхности органа (рис. 88, В).
В рыхлой колленхиме клетки на очень ранней стадии формирования разъединяются в углах с последующим образованием схизогенных межклетников; утолщение оболочек происходит на тех участках стенок, которые примыкают к межклетникам (рис. 88, Г). Рыхлую колленхиму часто называют колленхимой с межклетниками, она как бы соединяет признаки уголковой колленхимы с функцией проветривания тканей.
Колленхима широко распространена среди двудольных, у однодольных если и встречается, то обычно лишь в области стеблевых узлов соломины злаков, что предохраняет растения от полегания. Колленхима характерна для растущих травянистых стеблей, черешков листьев, цветоножек и т. д. Обычно она размещается в несколько слоев по периферии органа под эпидермисом. Изредка в механической обкладке жилок листьев некоторых двудольных встречаются клетки, напоминающие колленхиму. Обкладка из такой «колленхимы» играет роль упругой (пружинящей) арматуры. У древесных пород колленхиму можно наблюдать и у однолетних и у двулетних побегов до разрушения первичной коры, перед появлением перидермы. При этом колленхимные клетки постепенно смешиваются с клетками паренхимы коры.
Особую форму колленхимы представляет гиподерма листа (субэпидермальный слой клеток), выполняющая наряду с механической функцией функцию дополнительной защиты листа от излишнего испарения (например, в хвое сосны или ели).
СКЛЕРЕНХИМА
Склеренхима (греч. skleros — твердый) — наиболее важная механическая ткань, которая встречается в органах почти всех высших растений. Клетки ее имеют прозенхимную форму и представляют собой волокна, длинные, плотно расположенные, с заостренными концами и утолщенной, чаще всего одревесневшей оболочкой. Отдельные клетки склеренхимы носят название склеренхимных клеток или элементарных волокон, в совокупности же они образуют склеренхимную ткань. Отдельный тяж склеренхимных волокон часто называют техническим волокном. Многие авторы относят к склеренхиме также склереи-ды. Вообще существует множество систем, предложенных для классификации склеренхимной ткани.
Волокна встречаются в корне, стебле, листьях, плодах, цветках, в виде отдельных тяжей (пучков) или цилиндров (сплошное кольцо на поперечном срезе), группами или рассеянно в проводящей ткани. Существует несколько типов распределения волокон, специфичных для двудольных или однодольных.
По сравнению с колленхимой склеренхимные волокна отличаются большей упругостью, равной 15—20 кг/мм2, тогда как у колленхимы она составляет не более 10—12 кг/мм2. В связи с этим К. Эсау (1980) называет эти волокна эластичными (упруго растянутыми), колленхиму же — пластичной, так как даже при сравнительно малых нагрузках она остается необратимо растянутой, что имеет свои преимущества для растущих органов.
Наличие склеренхимы дает возможность органам растения противостоять нагрузкам, которые возникают в результате изгиба или под действием массы самого растения.
В отличие от клеток колленхимы у склеренхимы стенки клеток утолщены равномерно, часто одревесневают, но могут остаться и целлюлозными (у льна, например). Живое их содержимое, даже без одревеснения оболочек, обычно отмирает.
По происхождению различают первичную и вторичную склеренхиму. Первичная склеренхима дифференцируется из клеток основной меристемы апексов, прокамбиальных пучков или из пе-рициклических волокон. Вторичную склеренхиму формирует камбий. В зависимости от расположения в теле растения и функциональных особенностей волокна подразделяют на две большие группы — древесинные (ксилемные) волокна, входящие в состав проводящей ткани ксилемы (древесины, см. Ксилема), и волокна различных систем тканей вне ксилемы — экстраксилярные волокна. Часто экстраксилярные волокна объединяют в одну группу, носящую название лубяные волокна.
Лубяные волокна расположены обычно в коровой части осевого органа, во флоэме (лубе, см. дальше), они встречаются в коре стебля и корня, а также в листовых черешках и пластинках, в цветоножках, плодоножках, реже в плодах (рис. 89, I, А — В). Чаще они находятся в стеблях травянистых растений, но у многих пальм образуются в листьях (рис. 89, /, Г). Клетки лубяных волокон длинные, толстостенные. Длина их колеблется у разных видов: у льна она составляет в среднем 40—60 мм (у некоторых сортов до 120 мм), у кендыря — 2—55 мм, у крапивы — около 80 мм. Самые длинные волокна были найдены у субтропического прядильного растения рами (китайская крапива) — до 350—500 мм. У некоторых растений оболочки лубяных волокон остаются целлюлозными (лен, рами), но вторичные утолщения стенок волокна составляют до 70—90 % площади его поперечного сечения. У крапивы, а также у рас конопли оболочки лубяных волокон быстро одревесневают на всем протяжении стебля, характер одревеснения ра-соспецифичен (рис. 89, //, 1—6).
Особенность оболочек лубяных волокон хорошо изучена под электронным микроскопом. На ультратонких поперечных срезах при малом увеличении микроскопа оболочки обычно выглядят двухслойными: наружный слой, темный гомогенный и очень тонкий (0,2—0,3 мкм), составлен срединной пластинкой и первичной оболочкой смежных клеток, внутрь от него располагается более толстый (до нескольких микрометров) и светлый слой вторичной оболочки (В2), в которой может выявляться тончайшая концентрическая ламеллированность. У лубяных волокон льна к полости клетки часто прилегает узкий дополнительный слой (В3) с волнистым контуром, в кото-
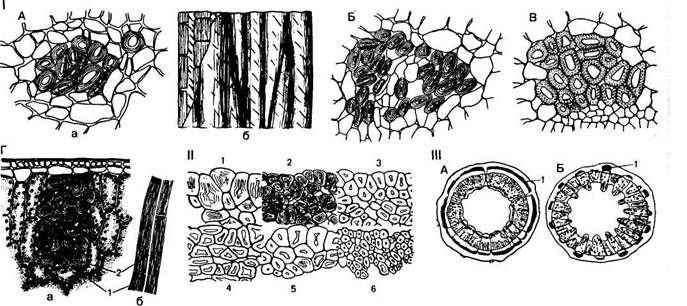
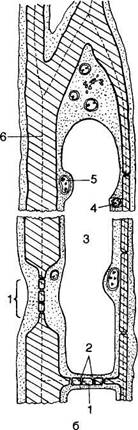
Рис. 89. Лубяные волокна растений:
Поперечные н продольные срезы лубяных волокон: Л — в стебле льна обыкновенного (Iinum uatatissum): а — поперечный разрез, б — продольный срез; Б — в стебле конопли посевной (Cannabis sativa); В — в черешке листа тополя канадского (Populus canadenas); Г — в лнегьях пальмы веерной (Trachycarpus excelsa): а — поперечный, б — продольный срезы; 1 — оболочка волокна; 2 — полоса волокна; II — разнообразие лубяных волокон у различных рас конопл (1—6); III — расположение лубяных волокон (Г): А — сплошным слоем в стебле рами; Б — отдельными пучками в стебле подсолнечника
ром также выявляется ламеллированность; незадолго до конца формирования вторичной оболочки цитоплазма распределена неравномерно по длине волокна: на некоторых участках она имеет вид очень узкого постенного слоя субмикроскопической толщины, в котором почти нет крупных органоидов и полость клетки занята в основном крупной вакуолью. В других участках цитоплазма образует скопления со многими органоидами, центральная вакуоль отсутствует. В целом волокна насыщены цитоплазмой, в которой встречаются единичные мелкие хлоропласты, часто с крахмальными зернами. Система гран в них развита слабо, хотя у некоторых насчитывается до 15 ти-лакоидов. В строме хлоропластов иногда выявляются пластидные рибосомы и пластоглобулы. Число митохондрий в несколько раз больше, чем хлоропластов, они содержат хорошо развитую систему крист. Характерная черта молодых лубяных волокон — высокоактивный комплекс Голь-джи. Довольно многочисленные диктиосомы продуцируют огромное число секреторных пузырьков, которые, очевидно, участвуют в формировании внутреннего слоя (В3) волокна. Элементы эндоплазматического ретикулума — в виде прозрачных пузырьков. У вполне сформировавшихся волокон протопласт чаще всего отмирает и полость клетки совершенно исчезает. У виноградной лозы, например, полость клетки разделе-на тонкими перегородками (перегородчатые лубяные волокна).
За счет интрузивного роста лубяных клеток создается исключительная прочность ткани, которая повышается благодаря спиральным расположениям микрофибрилл. Обычно лубяные волокна составляют простую ткань, располагаясь либо более или менее широким поясом, либо собраны отдельными группами, образующими вместе с проводящими тканями сосудисто-волокнистые пучки (рис. 89, ///, А, Б). У некоторых растений лубяные волокна с паренхимными клетками образуют лубяную паренхиму.
Свойства лубяных волокон (прочность, исключительная эластичность, большая длина волокна, отсутствие одревеснения) очень ценны для текстильной промышленности. Особый интерес представляют такие растения, как лен, конопля, кенаф, джут, рами. Они возделываются человеком уже более 5000 лет, их техническое волокно используется в текстильной промышленности.
Текстильные качества лубяных волокон неодинаковы. Различают твердо- и мягковолокни-стые растения. У мягковолокнистых (лен) волокна мягкие, эластичные, идут на изготовление высококачественных льняных тканей. У твердо-волокнистых (новозеландский лен, виды агавы, некоторые пальмы) лубяные волокна твердые, грубые, сравнительно малоэластичные. Из таких волокон делают веревки, щетки, грубые волокнистые ткани.
По происхождению лубяные волокна бывают первичными и вторичными. Первичные волокна образуются перициклом, вторичные — камбием. У травянистых двудольных растений преобладают первичные волокна, у древесных — вторичные.
Вторичные лубяные волокна намного короче первичных и чаще одревесневают. У конопли длина первичных волокон в среднем составляет 10— 12 мм, а вторичных — 2—3 мм. Вторичные волокна хорошо развиты у липы, березы, дуба и других древесных растений. Крайне редко лубяные волокна развиваются у голосеменных.
Древесинные волокна, или либриформ (лат. liber, род. п. libri — луб, лыко и forma — форма),—специализированные механические элементы древесины. Формируются они камбием, имеют, как правило, одревесневшие вторичные оболочки.
В эволюционном плане волокна либриформа произошли из элементов ксилемы, в которых функция проведения воды сочетается с опорной функцией, т. е., вероятнее всего, из трахеид (см. дальше). Свидетельство тому — наличие постепенных переходов между этими двумя типами клеток у некоторых покрытосеменных, например у дуба. Это родство подтверждается такими изменениями клеток древесинных волокон, как уменьшение длины волокна, утолщение стенок, уменьшение размеров пор и изменение формы их отверстий.
Волокна либриформа значительно короче лубяных волокон (не более 2 мм), одревесневшие оболочки снабжены простыми порами со щеле-видными очертаниями. Так же, как у лубяных волокон, имеющих простые поры, поры либриформа расположены по спирали.

|

|

|
| Рис. 90. Разнообразие древесных волокон: 1 — живые паренхима-газированные; 2, 3 — переходного тала; 4 — механическое волокно |
Либриформ нередко называют многофункциональной тканью (В. Г. Александров, 1966). Это связано с изменчивостью морфоструктуры волокон. Часто древесинные волокна морфологически выражены нечетко. Встречаются переходные элементы между клетками либриформа и во-допроводящими элементами. В этом случае клетки либриформа снабжены большим числом окаймленных пор и принимают участие в транспортировке воды. У таких древесинных волокон
рано отмирает живое содержимое. Нередко наблюдаются переходные элементы между клетками либриформа и древесинной паренхимой. В этом случае клетки волокон значительно укорочены, на их стенках имеется множество простых пор, они сохраняют живое содержимое и по многим особенностям напоминают живую паренхиму. В таких клетках запасаются крахмал и другие органические вещества.
У некоторых растений полость волокон либриформа перегорожена неполными тонкими перегородками. Такой либриформ называется перегородчатым, встречается он, например, в древесине винограда (рис. 90).
Клетки либриформа очень прочны, но почти неэластичны. Главная его функция — опора для водопроводящих тканей и для всего растения. Этот тип склеренхимы весьма распространен среди высших растений. У лиственных деревьев либриформ иногда занимает значительную часть древесины, особенно в тех ее массивах, которые сформировались во вторую половину вегетационного периода. Иногда склеренхима формируется перициклом, в этом случае волокна называют перициклическими. Они долго сохраняют целлюлозные стенки, редко одревесневают.
| <== предыдущая лекция | | | следующая лекция ==> |
| Суп картопляний з консервами з бобових | | | Желе рибне |