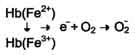В. Обезвреживание активных форм кислорода в эритроцитах
Большое содержание кислорода в эритроцитах определяет высокую скорость образования супероксидного анион-радикала (О2-), пероксида водорода (Н2О2) и гидроксил радикала (ОН.). Эритроциты содержат ферментативную систему, предотвращающую токсическое действие активных форм кислорода и разрушение мембран эритроцитов (рис. 14-4). Постоянный источник активных форм кислорода в эритроцитах - неферментативное окисление гемоглобина в метгемоглобин:
В течение суток до 3% гемоглобина может окисляться в метгемоглобин. Однако постоянно
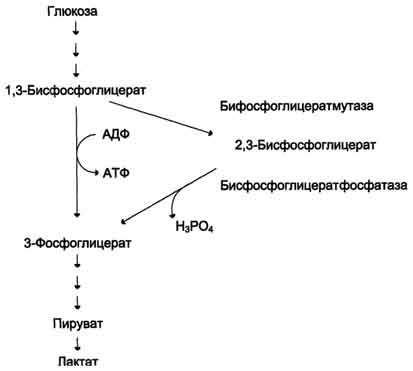
Рис. 14-3. Метаболизм 2,3-бисфосфоглицерата в эритроцитах. метгемоглобинредуктазная сисгема восстанавливает метгемоглобин в гемоглобин. Метгемоглобинредуктазная сисгема состоит из цитохрома B5 и флавопротеина цитохром B5редуктазы, донором водорода для которой служит NADH, образующийся в глицеральдегиддегидрогеназной реакции гликолиза (рис. 14-4). Цитохром B5 восстанавливает Fe3+ метгемог-лобина в Fe2+: Hb-Fe3+ + цит. b5 восст. → HbFe2+ + цит. b5 ок.. Окисленный цитохром B5 далее восстанавливается цитохром B5 редуктазой: Цит. B5 ок + NADH → цит. B5 восст. + NAD+. Супероксидный анион с помощью фермента супероксидцисмутазы превращается в пероксид водорода: O2- + O2- + Н+ → H2О2 + O2. Пероксид водорода разрушается каталазой и содержащим селен ферментом глутатионпероксидазой. Донором водорода в этой реакции служит глутатион - трипептид глутамилцистеинилглицин (GSH) (см. раздел 12). 2Н2О → 2Н2О + О2; 2GSH + 2Н2О2 → GSSG + 2Н2О. Окисленный глутатион (GSSG) восстанавливается NADPH-зависимой глутатионредуктазой. Восстановление NADP для этой реакции обеспечивают окислительные реакции пентозофосфатного пути (см. раздел 7).
|