Лекция №2 от 20 апреля 2002 г.
Важным фактором, определяющим продукцию гормона, является состояние регулируемой функции, т.е. выработка гормонов регулируется по принципу саморегуляции. Отечественный ученый М.М.Завадовский в 1941 году, изучая закономерности в регуляции деятельности эндокринных желез, впервые сформулировал принцип «плюс-минус взаимодействие», получивший в дальнейшем название «принцип обратной связи». Различают положительную обратную связь, когда повышение уровня гормона в крови стимулирует высвобождение другого гормона (повышение уровня эстрадиола вызывает высвобождение ЛГ в гипофизе), и отрицательную обратную связь, когда повышенный уровень одного гормона угнетает секрецию и высвобождение другого (повышение концентрации тиреоидных гормонов в крови снижает секрецию ТТГ в гипофизе). Вся гормональная регуляция осуществляется механизмами, функционирующими по принципу обратной связи, в которых четко выделяются различные уровни взаимодействия. Первый уровень (или контур) саморегуляции наиболее простой тип обратной связи. Здесь продукция гормона мало зависит от гипофиза и гипоталамуса, а в основном определяется изменением химического состава крови. Схематически это можно представить так:
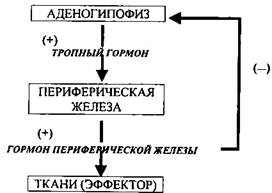
Второй контур саморегуляции замыкается на уровне гипофиза, который контролирует работу многих эндокринных желез. Этот механизм осуществляется за счет тропных гормонов. Например, гипофиз выбросил в кровь избыток ТТГ, щитовидная железа усиливает выработку тироксина и трийодтиронина, они в крови находятся в избытке (плюс взаимодействие). Повышенная концентрация гормона в крови автоматически уменьшает дальнейшую выработку ТТГ (минус взаимодействие). Таким образом, гипофиз включен в систему нервно-гуморальной регуляции, работающей по обратной гуморальной связи, которая автоматически поддерживает продукцию гормонов железами-мишенями. Схематически это выглядит так:
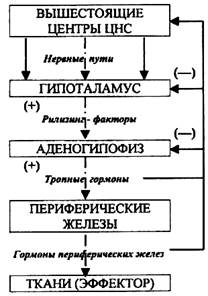
Наконец, имеется третий контур саморегуляции эндокринных желез, который замыкается на уровне гипоталамуса, входящего в состав промежуточного мозга. Некоторые ядра гипоталамуса обладают способностью к нейросекреции — выработке нейромедиаторов (дофамин, серотонин, норадреналин, ацетилхолин, γ‑аминомасляная кислота), принимающих участие в регуляции психической деятельности и поведения, а также нейрогормонов, регулирующих функциональную активность гипофиза. Нейросекреты — это промежуточное звено между нервными и гуморальными механизмами регуляции, которое соединяет их воедино. Гипоталамус и гипофиз имеют общее кровоснабжение. Кровь оттекающая от гипоталамуса через портальную систему омывает гипофиз и оказывает влияние на выработку им гормонов. Гипофизотропные гормоны гипоталамуса делятся на гормоны, усиливающие (высвобождающие, рилизинг‑гормоны, либерины) и угнетающие (ингибирующие, статины) выделение соответствующих тропных гормонов передней доли гипофиза.
Выработка нейросекрета гипоталамусом регулируется нервным путем, но главная роль в образовании этих нейрогормонов принадлежит механизму обратной гуморальной связи. Их продукция зависит от концентрации в крови гормонов той или иной железы и соответствующего гормона гипофиза, а также от информации поступающей от тканей, потребляющих данные гормоны. В регуляции эндокринных желез принимает участие и ЦНС. Большинство нервных и гуморальных путей регуляции сходится на уровне гипоталамуса и благодаря этому в организме образуется единая нейроэндокринная регуляторная система. В регуляции некоторых гормонов нервная система занимает ведущую роль. Например, выработка адреналина мозговым слоем надпочечников, главным образом, зависит от активности симпатической нервной системы. Поэтому при стрессовых ситуациях, когда возбуждается симпатический отдел ВНС, в крови резко повышается концентрация адреналина. Большое значение нервные влияния имеют и в регуляции выработки гормонов гипофиза. Так при болевом раздражении увеличивается количество АКТГ. Но и характер процессов, протекающих в ЦНС, во многом определяется состоянием эндокринной регуляции. Нервная система, эволюционно более поздняя, имеет как управляющие, так и подчиненные связи с эндокринной системой. Эти две регуляторные системы дополняют друг друга, они образуют функционально единый механизм, обеспечивающий согласованность всех процессов жизнедеятельности. Вещества, которые выделяются из пресинаптических нервных окончаний в синаптическую щель и вызывают биологический эффект, связываясь с рецепторами постсинаптической мембраны, называют нейромедиаторами.
[1] +601+С.99-100 [2] ++601+С.99 [3] ++601+С.101 [4] ++601+С.101 [5] ++601+С.101 [6] ++421+С.131 [7] ++421+С.131 [8] ++601+ С.243 [9] ++601+С.101 [10] нейроэндокринную, используя в качестве средств управления нейрогормоны. [11] Одним из простейших вариантов является изменение деятельности клеток под влиянием продуктов обмена веществ. Последние могут изменять работу клетки, из которой происходит выделение этих продуктов, и других органов, расположенных на достаточном удалении. Например, под влиянием СО2, образующегося в тканях в результате утилизации кислорода, изменяется активность центра дыхания и как следствие — глубина и частота дыхания. Под влиянием адреналина, выделяемого в кровь из надпочечников, изменяются частота и сила сердечных сокращений, тонус периферических сосудов, ряд функций центральной нервной системы, интенсивность обменных процессов в скелетных мышцах, увеличиваются коагуляционные свойства крови. [12] ++750+С.521 [13] ++743+С.261 [14] 10бр [15] ++638+С.215, ++745+С.5-6 [16] --145- С.353 [17] аспекты ++601+С.99
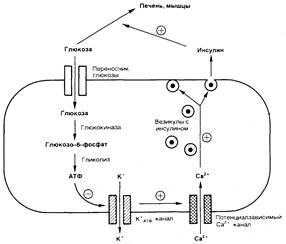
Лекция №2 от 20 апреля 2002 г. ГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ Функциональная активность эндокринной железы может регулироваться «субстратом», на который направлено действие гормона. Так, глюкоза стимулирует секрецию инсулина из b ‑клеток панкреатических островков (островки Лангерганса), а инсулин понижает концентрацию глюкозы в крови, активируя ее транспорт в мышцы и печень. Это происходит следующим образом. Глюкоза входит в b ‑клетки поджелудочной железы через переносчик глюкозы и сразу же фосфорилируется глюкокиназой, после чего вовлекается в гликолиз. Образующийся при этом АТФ ингибирует калиевые каналы, вследствие чего снижается мембранный потенциал b ‑клеток и активируются потенциалзависимые кальциевые каналы. Входящий в b ‑клетку кальций стимулирует слияние везикул, содержащих инсулин, с плазматической мембраной и секрецию инсулина. Инсулин активирует перенос глюкозы в печень, сердце и скелетные мышцы, вследствие чего уровень глюкозы в крови снижается, замедляется ее вход в b ‑клетки и уменьшается секреция инсулина (рис. 5.1).
Рис. 5.1. Регуляция уровня гормона в крови по механизму обратной связи. Объяснение в тексте. Такой же механизм лежит в основе секреции паратгормона (паратиреоидный гормон, паратирин) и кальцитонина. Оба гормона влияют на концентрацию кальция и фосфатов в крови. Паратиреоидный гормон вызывает выход минеральных веществ из кости и стимулирует реабсорбцию кальция в почках и кишечнике, в результате чего возрастает концентрация кальция в плазме крови. Кальцитонин, напротив, стимулирует поступление кальция и фосфатов в костную ткань, в результате чего концентрация минеральных веществ в крови снижается. При высокой концентрации кальция[V.G.1] в крови подавляется секреция паратиреоидного гормона и стимулируется секреция кальцитонина. В случае снижения концентрации кальция в крови секреция паратиреоидного гормона усиливается, а кальцитонина — ослабляется. Такая регуляция постоянства внутренней среды организма, происходящая по принципу отрицательной обратной связи, очень эффективна для поддержания гомеостаза, однако не может выполнять все задачи адаптации организма. Например, кора надпочечников продуцирует стероидные гормоны в ответ на голод, болезнь, эмоциональное возбуждение и т.п. Чтобы эндокринная система могла «отвечать» на свет, звуки, запахи, эмоции и т.д., должна существовать связь между эндокринными железами и нервной системой. Основные связи между нервной и эндокринной системами регуляции осуществляются посредством взаимодействия гипоталамуса и гипофиза (рис. 5.2). Нервные импульсы, приходящие в гипоталамус, активируют секрецию так называемых рилизинг‑факторов (либеринов и статинов): тиреолиберина, соматолиберина, пролактолиберина, гонадолиберина и кортиколиберина, а также соматостатина и пролактостатина. Мишенью для либеринов и статинов[V.G.2], секретируемых гипоталамусом, является гипофиз. Каждый из либерияов взаимодействует с определенной популяцией клеток гипофиза и вызывает в них синтез соответствующих тропинов: тиреотропина, соматотропного гормона (соматотропин — гормон роста), пролактина, гонадотропного гормона, (гонадотропины — лютеинизирующий и фолликулостимулирующий), а также адренокортикотропного гормона (АКТГ, кортикотропин). Статины оказывают на гипофиз влияние, противоположное действию либеринов, — подавляют секрецию тропинов. Тропины, секретируемые гипофизом, поступают в общий кровоток и, попадая на соответствующие железы, активируют в них секреторные процессы.
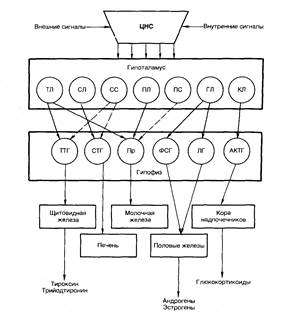
Рис. 5.2. Регуляция активности эндокринных желез центральной нервной системой при участии гипоталамуса и гипофиза. ТЛ — тиреолиберин; СЛ — соматолибсрин; СС — соматостатин; ПЛ — пролактолибсрии; ГЛ — гонадолибсрин; КЛ — кортиколибсрин; ТТГ — тирсотропный гормон; СТГ — соматотропный гормон (гормон роста); Пр — пролактин; ФСГ — фолликулостимулирующий гормон; ЛГ — лютеинизирующий гормон; АКТГ — адренокортикотропный гормон. Сплошными стрелками обозначено активирующее, пунктирными — ингибирующсе влияние.
Молекула соматолиберина является самой крупной среди либеринов, она состоит из 15 аминокислотных остатков; самая маленькая[V.G.3] молекула — трипептид — у тиреолиберина. Молекулы тропинов, образующихся в гипофизе, содержат от 13 до 198 аминокислотных остатков.
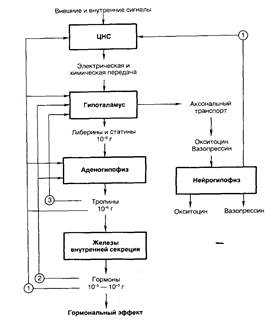
Рис. 5.3. Прямые и обратные связи в нейроэндокринной системе регуляции. 1 — медленно развивающееся и продолжительное ингибирование секреции гормонов и нейромедиаторов, а также изменение поведения и формирование памяти; 2 — быстро развивающееся, но продолжительное ингибирование; 3 — кратковременное ингибирование. Регуляция деятельности гипофиза и гипоталамуса, кроме сигналов, идущих «сверху вниз», осуществляется гормонами «исполнительных» желез (рис. 5.3). Эти «обратные» сигналы поступают в гипоталамус и затем передаются в гипофиз, что приводит к изменению секреции соответствующих тропинов. После удаления или атрофии эндокринной железы стимулируется секреция соответствующего тропного гормона; при гиперфункции железы секреция соответствующего тропина подавляется. Обратные связи не только позволяют регулировать концентрацию гормонов в крови, но и участвуют в дифференцировке гипоталамуса в онтогенезе. Образование половых гормонов в женском организме происходит циклически, что объясняется циклической секрецией гонадотропных гормонов. Синтез этих гормонов контролируется гипоталамусом, образующим рилизинг-фактор этих тропинов (гона[V.G.4] долиберин). Если самке пересадить гипофиз самца, то пересаженный гипофиз начинает функционировать циклично. Половая дифференцировка гипоталамуса происходит под действием андрогенов. Если самца лишить половых желез, продуцирующих андрогены, то гипоталамус будет дифференцироваться по женскому типу. В железах внутренней секреции иннервированы, как правило, только сосуды, а эндокринные клетки изменяют свою биосинтетическую и секреторную активность лишь под действием метаболитов, кофакторов и гормонов, причем не только гипофизарных. Так, ангиотензин II стимулирует синтез и секрецию альдостерона. Отметим также, что некоторые гормоны гипоталамуса и гипофиза могут образовываться не только в этих тканях. Например, соматостатин (гормон гипоталамуса, ингибирующий образование и секрецию гормона роста) обнаружен также в поджелудочной железе, где он подавляет секрецию инсулина и глюкагона. Большинство нервных и гуморальных путей регуляции сходится на уровне гипоталамуса и благодаря этому в организме образуется единая нейроэндокринная регуляторная система. К клеткам гипоталамуса подходят аксоны нейронов, расположенных в коре больших полушарий и подкорковых образованиях. Эти аксоны секретируют различные нейромедиаторы, оказывающие на секреторную активность гипоталамуса как активирующее, так и тормозное влияние. Поступающие из мозга нервные импульсы гипоталамус «превращает» в эндокринные стимулы, которые могут быть усилены или ослаблены в зависимости от гуморальных сигналов, поступающих в гипоталамус от желез и тканей, подчиненных ему. Тропины, образующиеся в гипофизе, не только регулируют деятельность подчиненных желез, но и выполняют самостоятельные эндокринные функции. Например, пролактин оказывает лактогенное действие, а также тормозит процессы дифференцировки клеток, повышает чувствительность половых желез к гонадотропинам, стимулирует родительский инстинкт. Кортикотропин является не только стимулятором стероидогенеза, но и активатором липолиза в жировой ткани, а также важнейшим участником процесса превращения в мозге кратковременной памяти в долговременную. Гормон роста может стимулировать активность иммунной системы, обмен липидов, сахаров и т.д. В задней доле гипофиза (нейрогипофиз) депонируются антидиуретический гормон (вазопрессин) и окситоцин (см. рис. 5.3). Первый вызывает задержку воды в организме и повышает тонус сосудов, второй стимулирует сокращение матки при родах и секрецию молока. Оба гормона синтезируются в гипоталамусе, затем транспортируются по аксонам в заднюю долю гипофиза, где депонируются и потом секретируются в кровь. Характер процессов, протекающих в ЦНС, во многом определяется состоянием эндокринной регуляции. Так, андрогены и эстрогены формируют половой инстинкт, многие поведенческие реакции. Очевидно, что нейроны, точно так же как и другие клетки нашего организма, находятся под контролем гуморальной системы регуля[V.G.5] ции. Нервная система, эволюционно более поздняя, имеет как управляющие, так и подчиненные связи с эндокринной системой. Эти две регуляторные системы дополняют друг друга, образуют функционально единый механизм, что обеспечивает высокую эффективность нейрогуморальной регуляции, ставит ее во главе систем, согласующих все процессы жизнедеятельности в многоклеточном организме.
|