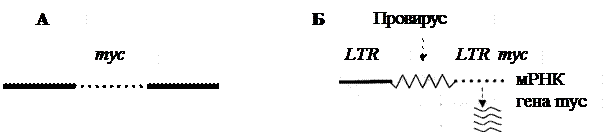Механизмы превращения протоонкогена в онкоген
Превращение протоонкогена в активно действующий онкоген обеспечивается следующими механизмами. 1. Присоединение к протоонокгену промотора – участка ДНК, с которым связывается РНК-полимераза, инициирующая транскрипцию гена, в том числе и онкогена, располагающегося непосредственно за ним (рис.1). Такого рода участки (промоторы) содержатся в больших терминальных повторах (LTR) ДНК-копий РНК-содержащих вирусов. Роль промотора могут выполнять и транспозирующие элементы генома – мобильные генетические элементы, способные перемещаться по геному и встраиваться в различные его участки.
Рис. 1. Схема процесса активации протоонкогена в результате вставки промотора (по R.K. Marray et al.): неактивный в нормальной хромосоме кур ген туc (А), оказавшись рядом (Б), вслед за LTR провируса лейкоза птиц после его интеграции в геном клетки активируется
2. Вставка в геном клетки энхансера (enchancer – усилитель) – участка ДНК, способного активизировать работу структурного гена, находящегося не только в непосредственной близости от него, но и на расстоянии многих тысяч пар нуклеотидов или даже встроенного в хромосому после него. Свойствами усилителя обладают подвижные гены, LTR ДНК-копий. В случае, проиллюстрированном рис. 2, LTR провируса не может работать как промотор (см. рис. 1, Б) и выступает в роли энхансера, в результате чего ген тус активизируется и транскрибируется.
Рис. 2. Схема процесса активизации протоонкогена в результате вставки энхансера (по R.K. Marray et al., 1988): А –нормальная хромосома кур, содержащая неактивный ген,mуc; Б –встраивание вируса лейкоза птиц в хромосому (в форме провируса) за myc-геном; активация и транскрибция гена туc.
3. Хромосомные абберации с явлениями транслокации,роль которых в механизмах опухолевой трансформации клетки можно проиллюстрировать следующим примером. При лимфоме Беркитта конец q-плеча хромосомы 8, отделившись от нее, переходит к хромосоме 14: гомологичный фрагмент последней перемещается к хромосоме 8; а неактивный ген туc (протоонкоген), находившийся в том ее сегменте, который попадает на хромосому 14, встраивается вслед за активными генами, кодирующими тяжелые цепи молекул иммуноглобулинов, и активизируется (рис. 3, 4). Явления реципрокной транслокации между 9-й и 22-й хромосомами имеют место в 95 % случаев миелоцитарного лейкоза. Хромосома 22 с укороченным в результате такой транслокации одним плечом получила название Филадельфийской.
Рис. 3. Схема реципрокной транслокации в клетках лимфомы Беркитта (пояснение в тексте)
Рис. 4. Схема процесса активизации mус-протоонкогена при транслокации в клетках лимфомы Беркитта (по R.K. Муггау, 1988): А – сегмент хромосомы 14 перед транслокацией, содержащий гены, кодирующие синтез иммуноглобулинов; Б – первично неактивный ген туc после транслокации оказывается под контролем энхансера, локализующегося в области генов, которые кодируют тяжелые цепи иммуноглобулина. В результате ген туc активизируется.
4. Точечные мутации протоонкогена,к примеру, C-H-raS, согласно 5. Амплификация (умножение) протоонкогенов,обладающих в норме 6. Трансдукция неактивных клеточных генов (протоонкогенов) в геном ретровируса и последующее их возвращение в клетку:считается, что онкоген опухолеродного вируса клеточного происхождения; при инфицировании животных или человека таким вирусом «похищенный» им ген попадает в иной участок генома, что и обеспечивает активизацию некогда «молчавшего» гена.
|