
Билет №27. 1.Формирование хромосомной теории
1. Формирование хромосомной теории В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911—1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности. Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов. В экспериментах он использовал плодовую мушку-дрозо-филу, обладающую важными для генетических экспериментов качествами: неприхотливостью, плодовитостью, небольшим количеством хромосом (четыре пары), множеством четко выраженных альтернативных признаков. Морган и его ученики установили следующее: 1. Гены, расположенные в одной хромосоме, наследуются совместно или сцепленно. 2. Группы генов, расположенных в одной хромосоме, образуют группы сцепления. Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и п+1 у гетерогаметных особей. 3. Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссин-говера возникают гаметы, хромосомы которых содержат новые комбинации генов. 4. Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера. За единицу расстояния между генами принимают 1 морганиду (1% кроссинговера) или процент появления кроссоверных особей. При значении этой величины в 10 морганид можно утверждать, что частота перекреста хромосом в точках расположения данных генов равна 10% и что в 10% потомства будут выявлены новые генетические комбинации. Основные положения хромосомной теории наследственности: • каждый ген имеет в хромосоме определенный локус (место); • гены в хромосоме расположены в определенной последовательности; • гены одной хромосомы сцеплены, поэтому наследуются преимущественно вместе; • частота кроссинговера между генами равна расстоянию между ними; • набор хромосом в клетках данного типа (кариотип) является характерной особенностью вида.
Для выяснения характера расположения генов в хромосомах и определения частоты кроссинговера между ними строят генетические карты. Карта отражает порядок расположения генов в хромосоме и расстояние между генами одной хромосомы. Эти выводы Моргана и его сотрудников получили название хромосомной теории наследственности. Важнейшими следствиями этой теории являются современные представления о гене как о функциональной единице наследственности, его делимости и способности к взаимодействию с другими генами.
Пример сцепленного наследования: Vg — нормальные крылья дрозофилы; vg — зачаточные крылья; ВВ — серая окраска тела; bb — темная окраска тела. Запись в хромосомном выражении:
В данном случае правило единообразия гибридов первого поколения соблюдается. В соответствии со вторым и третьим законами Менделя следовало ожидать при последующем анализирующем скрещивании по 25% каждого из возможных фенотипов (серых, длиннокрылых мух, серых короткокрылых, черных длиннокрылых и черных коротко-крылых). Однако опыты Моргана не дали таких результатов. При скрещивании рецессивной по обоим признакам самки VgVgbb с гибридным самцом из F1 образовалось 50% серых мух с короткими крыльями и 50% мух с черным телом и длинными крыльями:
Если же скрещивают дигибридную самку с гомозиготным рецессивным самцом, то в образуется потомство: 41,5% — серых с короткими крыльями, 41,5% — черных с длинными крыльями, 8,5% — серых с длинными крыльями, 8,5% — черных с короткими крыльями.
Данные результаты свидетельствуют о наличии сцепления генов и кроссинговере между ними. Так как в потомстве от второго скрещивания было получено 17% рекомбинант-ных особей, то расстояние между генами Vg и В равно 17%, или 17 морганидам.
Сцепленное наследование. Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10,4 и 23 пары соответственно. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе. Совместное наследование генов X Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному набору хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигибрид Было установлено, однако, что кроме обычных гамет возникают и другие — Аb и аВ — с новыми комбинациями генов, отличающимися от родительской гаметы. Причиной возникновения новых гамет является обмен участками гомологичных хромосом, или кроссинговер. Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных. Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам. Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная величина кроссинговера не превышает 50%. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования. Генетической картой хромосом называют схему взаимного расположения генов, находящихся в одной группе сцепления. Такие карты составляются для каждой пары гомологичных хромосом. Возможность подобного картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов. Цитологические карты хромосом, схематическое изображение хромосом с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Ц. к. х. составляют для организмов, для которых обычно уже имеются генетические карты хромосом. Каждое место расположения гена (локус) на генетической карте организма, установленное на основе частоты перекреста участков хромосом (кроссинговера), на Ц. к. х. привязано к определённому, реально существующему участку хромосомы, что служит одним из основных доказательств хромосомной теории наследственности. Для построения Ц. к. х. используют данные анализа хромосомных перестроек (вставки, делеции и др.) и, сопоставляя изменения морфологических признаков хромосом при этих перестройках с изменениями генетических свойств организма, устанавливают место того или иного гена в хромосоме. Пользуясь методом хромосомных перестроек, амер. генетик К. Бриджес составил в 1935 подробную Ц. к. х. плодовой мушки дрозофилы, наиболее полно генетически изученного организма. Гигантские хромосомы насекомых отряда двукрылых оказались самыми удобными для построения Ц. к. х., т.к. наряду с большими размерами обладают чёткой морфологической очерченностью: каждый участок этих хромосом имеет свой определённый и чёткий рисунок, обусловленный характерным чередованием по длине ярко окрашиваемых участков (дисков) и слабо окрашиваемых (междисков). Цитологическими методами легко определить отсутствие участка хромосомы или перенос его в др. место. Сопоставление Ц. к. х. с генетическими показало, что физическое расстояние между генами в хромосомах не соответствует генетическому (видимо, частота кроссинговера неодинакова в разных участках хромосом), поэтому плотность распределения генов на цитологических и генетических картах хромосом различна. Так было установлено важное генетическое явление - неравномерность частот перекреста по длине хромосомы. Линейное расположение генов и их последовательность, установленные генетическими методами, подтверждаются Ц. к. х. Современные методы цитологии и генетики позволяют построить Ц. к. х. многих организмов, в том числе человека. 2. Антропогенез, или происхождение человека, рассматривается как становление вида Человек разумный в процессе формирования общества (социогенеза). Краеугольный камень учения об антропогенезе - общая теория эволюции, к крупнейшим достижениям которой относится сформулированная впервые Ч. Дарвином симиальная гипотеза происхождения человека от высокоразвитых обезьян третичного периода, аргументация которой значительно расширилась и пополнилась многими новыми фактами из области сравнительной биохимии, иммунологии, этологии приматов, молекулярной антропологии и др. Эти материалы полностью подтвердили наибольшую близость человека к африканским человекообразным обезьянам (понгидам) и в первую очередь к шимпанзе. Антропогенез условно можно разделить на 5 стадий. 1. Понгидно-гоминидная стадия, стадия развития общей понгидно-гоминидной ветви, которая началась 16-18 млн лет и завершилась не позднее 6,0 млн лет тому назад разделением на 2 ветви: понгидную и гоминидную. 3. Архантропная стадия (2,0-0,5 млн лет тому назад). 4. Палеоантропная стадия (0,5-0,035 млн лет тому назад). 5. Неоантропная стадия (началась около 0,2 млн лет тому назад и длится по настоящее время). Ранний период эволюции общего понгидно-гоминидного ствола был представлен североафриканским олигоценовым египтопитеком и группой дриопитеков, в первую очередь древнейшим (нижнемиоценовым) дриопитеком африканским из Кении, возраст находок которого определяется примерно в 20 млн лет. На второй (прегоминидной) стадии около 5 млн лет назад появились двуногие человекообезьяны - австралопитековые, которые довольно широко распространились по Африке, а возможно, и за её пределами в период от 4,5 до 1 млн лет назад. Австралопитековые - это подсемейство вымерших южных человекообразных обезьян, включаемых в семейство гоминид. Рост взрослого австралопитека, передвигавшегося на двух ногах, не превышал 130 см, вес - 30-40 кг, объём мозговой полости достигал 700 см3. Лицевой череп и челюсти австралопитека многими признаками более напоминали таковые человека, чем приматов В одной из периферийных популяций ранних австралопитековых могли сформироваться первые представители рода Homo, сосуществовавшие с австралопитековыми на большом временном промежутке, что доказывается находками в Кении и Танзании. Существует также мнение о независимом происхождении австралопитековых и Homo habilis при конвергентном развитии двуногого хождения в обеих линиях. Однако только представители рода Homo были способны изготовлять орудия. К австралопитековым относят также более поздние находки - президжантропа, зиджантропа, па-рантропа, плезиантропа. Наиболее прогрессивные австралопитековые - президжантропы перейти к изготовлению орудий, создав самую раннюю олдовайскую (или галечную) культуру, и стали наиболее ранними представителями Человека умелого - Homo habilis. Ископаемые остатки ранних Homo (древностью около 2 млн лет) обнаружены в ряде районов Восточной Африки (Танзания, Кения, Эфиопия), нередко с праорудиями древнейшей олдовайской культуры человечества. Около 1,5 млн лет тому назад первый вид человека - Человек умелый (Homo habilis) сменился другим видом - Человек прямоходящий (Homo erectus). В дальнейшем, происходило постепенное расширение экологической ниши Homo, поглотившей экологические ниши австралопитековых. Ещё 1,3-1,4 млн. лет тому назад Homo erectus сосуществовал с поздними австралопитековыми, а приблизительно 1 млн. лет тому назад Homo erectus стал единственным представителем гоминид на Земле. Этот вид обнаруживается в различных регионах Африки и Евразии примерно до периода 0,3 млн лет. Ранние этапы гоминизации связаны с Африкой (системой Восточно-Африканского рифта), данный факт во многом объясняется тем, что это был гигантский горный разлом, изобиловавший водными источниками и разнообразными животными. В 1891 году французский врач Э. Дюбуа нашёл остатки представителя древних гоминид на острове Ява, которого назвал питекантропом. Он был ростом 170 см и передвигался на двух ногах, слегка согнутых в коленях. Питекантроп имел объём мозговой полости 900-1000 см3, небольшую выпуклость лба, массивные надглазничные валики, плоские носовые кости, тяжёлую нижнюю челюсть без подбородочного выступа. Во многом он был сходен с найденным в 1927 году близ Пекина синантропом с объёмом мозговой полости 1050 см3 По мнению многих эти две находки являются двумя географическими расами или двумя подвидами Homo erectus, представляющими третью (архантропную) стадию антропогенеза. Объём мозга составлял у них около 1000 см3, укороченные челюсти и расширенный альвеолярный отросток обеспечивали относительно большую подвижность языка, что даёт основания допускать возможность появления примитивной речи на архантропной стадии эволюции гоминид. Четвёртая (палеоантропная) стадия антропогенеза представлена неандертальцем (Homo sapiens neanderthalensis), первый экземпляр которого найден в 1856 году в долине реки Неандер (Германия). От современного человека он отличался прежде всего низким сводом черепа, покатым лбом и тяжёлым сплошным надглазничным валиком,, пяткообраз-но нависающим затылком, к которому прикреплялись сильно развитые мышцы шеи, недоразвитием подбородочного выступа и отклонением назад челюстного симфиоза Объём мозга неандертальца составлял в среднем 1450 см, несколько превышая объём мозга современного человека (1350 см3). Примерно 200-50 тыс. лет тому назад антропогенез вступил в последнюю (неоантропную) стадию. Самые ранние находки современного человека - кроманьонца (Homo sapiens sapiens) датируются в 40 тыс. лет (первая находка сделана во Франции близ деревни Кроманьон в 1868 году). Кроманьонцы отличались высоким ростом (до 180-190 см), обладали объёмом черепа, сравнимым с таковым современного человека, узким носом, широким прямым лбом и нижней челюстью с большим подбородочным выступом. Кроманьонцы искусно делали каменные и костяные орудия (рис. 169), в их пещерах найдены рисунки животных и сцен охоты. Кроманьонцы создали различные археологические культуры позднего палеолита. Такие качества неоантропов, как коллективная трудовая деятельность, развитая речь, интеллект, привели в процессе дальнейшего развития к возникновению человеческого общества. Важным этапом его развития явилась революция неолита (позднего каменного века), в результате которой человек одомашнил животных, окультурил растения и вышел из полной зависимости от природы. Благодаря этому он избежал гибели от голода в результате первой экологической катастрофы, когда истребил всех диких животных в позднем мезолите. Эволюция гоминид не была линейным процессом. Вероятны значительные вариации скоростей изменения в различных линиях и в одном и том же филетическом ряду в разное время. Можно полагать, что развитие гоминид шло на каждом этапе по нескольким направлениям (рис.170) и в пределах каждого направления возникали разновидности, которые изменялись под влиянием разных факторов эволюции.
3.
|
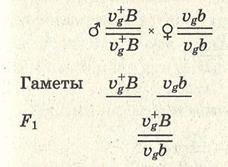


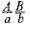 образует четыре типа гамет (АВ, Аb, аВ и аb) в равных количествах, то такой же дигибрид
образует четыре типа гамет (АВ, Аb, аВ и аb) в равных количествах, то такой же дигибрид 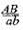 образует только два типа гамет: (АВ и аb) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
образует только два типа гамет: (АВ и аb) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.


