Модель оперона для управления генами. Lac-оперон.
Схема управл-я бактер-ми генами вперв. была предложена Франсуа Жакобом и Жаком Моно, использ-ть в кач-ве примера гены лактозы. Жакоб и Моно дали название различным элементам этой системы: репрессор, оператор, и т.д. Мн. бакт-ые гены объединены в группу, кот. транскрибируется вместе из одной и той же начальной точки, образуя одного РНК-посредника. Кластер генов, включенных вместе при транскрибировании из одного и того же промотора, назыв. опероном. (рисунок) Составные части оперона.
Несмотря на то, что высшие организмы имеют больше генов, чем бакт., они не имеют оперонов. Их гены регулируются по одному. Тем не менее, гены высш. орг-ов регулируются путем присоедин-я контролир-х б-ов, как глобальных, так и специфических, перед геном. Некот-е опероны имеют только один ген, больш-во имеют от 2х до полудюжины и немногие имеют больше. Например, lac-оперон, кот-ый включает 3 гена: lac Z, lac Y, lac A. Включен lac-оперон или выкл-ен зависит от 2х регуляторных б-ов: lac L и Crp. Только когда репрессор lac L отсутствует и б-к Crp присутствует в кач-ве вспомагат-ого, РНК-полимераза способна присоединятся к промотору м создать РНК- посредника. Lac-оперон: включен или выкл.(рисунок)
РНК-посредник транскрибируется с помощью только одной нити ДНК, выступающей в качестве матричной нити. Вторая нить ДНК не используется. Но, предположим, что мы все же использовали нематричную нить и транкрибировали из нее РНК. При этом должна получиться мол-ла РНК, комплиментарная по последовательности РНК-посреднику. Эта РНК называется антисмысловой-РНК, и она может образовывать пару со своей комплиментарной РНК-посредником также как две нити ДНК в исходном гене образуют пару друг с другом. Антисмысловая-РНК время от времени используется бактериями и высшими организмами в регуляции генов. Если содаётся антисмысловая-РНК, она образует пару с РНК-посредником и препятсвует ее присоединению к рибосоме. Соответственно, РНК-посредник не может участвовать в синтезе белка и данный ген отключается, даже если была образована РНК-посредник. На практике антисмысловая-РНК не образуется путем транскрибирования нематричной нити того же гена, что и РНК-посредник. Для создания анлисмысловой-РНК используется другой, отличный от данного «антиген». Бактериоферритин – белок, используемый бактериями для хранения лишних атомов железа. Ген bfr кодирует антисмысловую-РНК. Поскольку для блокировки РНК-посредника требуется всего лишь относительно короткая часть антисмысловой РНК, то последовательность антигена сходна с последовательностью исходного гена, однако она короче. Когда концентрация железа в культуральной среде невелика, бактериоферритин не требуется, но он образуется, если уровень содержания железа повышается. Сам по себе ген bfr транскрибируется для образования РНК-посредника в обоих случаях. Тем не менее, антиген bfr транскрибируется для создания антисмысловой РНК только при пониженном содержании железа. Это предотвращает синтез белка бактериоферритина при дефиците железа. Итак, чтоже вкл и выкл антиген bfr? Глобальный регуляторный белок, который называется «регулятор поглощения железа» (ISP), обнаруживает и присоединяет железо. Если имеется большое кол-во железа, ISP выступает в роли репрессора и прекращает транскрипцию дюжины или более оперонов, необходимых для адаптпции данной клетки к нехватке железа. В частности, ISP+железо откл ген anti-bfr, в результате чего начинается выработка бактериоферритина. Т.о., используя антисмысловую РНК, мы можем регулировать один ген в противоположность регулированию ряда других. Искуственно синтезированная антисмысловая РНК препятствует экспрессии генов или какому-либо другому процессу с участием РНК. Антисмысловая-РНК в настоящее время проходит эксперементальные испытания на способность задерживать развитие рака путем остановки деления хромосом.
В эукариотических клетках присутствуют три ядерные РНК-полимеразы – Ι, ΙΙ и ΙΙΙ. РНК-полимеразаΙ находится в ядрышке и участвует главным образом в биосинтезе рРНК, в то время как РНК-полимеразы ΙΙ и ΙΙΙ обнаруживаются в хроматине и нуклеоплазме. РНК-полимераза ΙΙ осуществляет синтез мРНК, РНК-полимераза ΙΙΙ отвечает за синтез тРНК, и 5S-рРНК. Митохондрии, подобно бактериям, содержат только одну РНК-полимеразу. S- это коф-т седементации (осаждения) при центрифугировании, его принято выражать в единицах Сведберга. Удлинение цепи РНК с помощью эукариотических ДНК-зависимых РНК-полимераз происходит таким же образом, как и при участии фермента из Е.соli, однако эти ферменты отличаются друг от друга по своей субъединичной стр-ре и по регуляторным элементам. Это, естественно, поскольку эукариотические РНК-полимеразы должны транскрибировать ДНК, которая плотно упакована в нуклеосомы и связана с другими ядерными белками хроматина.
ДНК-зависимую РНК-полимеразу можно избирательно ингибировать. Элонгация цепей РНК с помощью РНК-полимеразы специфически ингибируется а/б актиномицином D как у прокариот, так и у эукариот. Плоская часть молекулы этого а/б интеркалирует в двойную спираль ДНК между соседними парами G-C деформируя матрицу ДНК. Эта локальная деформация мешает движению полимеразы вдоль матрицы. Т.о., актиномицин D вызывает как бы «заедание молнии». Другим интеркалирующим ингибитором является акридин, мол-лы которго также имеют плоскую стр-ру. Ещё одним важным а/б, ингибирующим синтез РНК, является рифампицин, который связывается с β-субъединицей прокариотической РНК-полимеразы и препятсвует инициации синтеза РНК. Однако у эукариот рифампицин не ингибирует синтез РНК. Специфическое ингибирующее действие на синтез РНК в животных клетках клетках оказывает α-аманитин – токсическое начало ядовитого гриба Amanita phalloides. Этот ингибитор блокирует синтез мРНК, осуществляемый эукариотической РНК-полимеразой ΙΙ, но не влияет на синтез РНК у прокариот. Т.о. Amanita phalloides вырабатывает в-во, нарушающее синтез РНК в прочих эукариотических организмах, но являющегося безвредным для его собственного транскрипционного аппарата.
Транкрипты РНК, синтезированные РНК-полимеразой, обычно претерпевают дальнейшие ферментативные превращения, называемые посттранскрипционным процессингом, и только после этого они обретают свою функциональную активность. рРНК и тРНК синтезируются в виде более длинных предшественников, которые затем модифицируются и расщепляются с образованием конечных продуктов. Транскрипты эукариотических мРНК также подвергаются процессингу, тогда как у прокариотических мРНК он отсутствует. рРНК как эукариотических, так и прокариотических клеток образуется из более длинных молекул-предшественников, называемых прерибосомными РНК. У прокариот 16S- и 23S-рРНК образуются из одного длинного 30S-предшественника. Этот предшественник метилируется по специфическим основаниям и расщепляется, давая 17S- и 25S- промежуточные РНК, которые затем процессируются путем отщепления остатков с помощью нуклеаз, образуя характерные для прокариот 16S- и 23S- рРНК, 5S-рРНК образуется отдельно из 3'-концевого участка 30S- предшественника. У эукариот 18S- и 28S-рРНК образуются в несколько этапов из большой 45S-прерибосомной РНК. Процессинг 45S-РНК протекает в ядрышке. Сначала происходит метилирование более чем 100 из 14000 нуклеотидов 45S-предшественника, модификации подвергаются главным образом 2'-гидроксильные группы рибозных остатков. Метилированная 45S-РНК претерпевает затем ряд ферментативных расщеплений, приводящих в конечном итоге к появлению 18S- и 5,8S-РНК, характерных для эукариотических рибосом. 5S-рРНК эукариот синтезируется отдельно. тРНК также образуются из более длинных РНК-предшественников в результате ферментативного удаления лишних нуклеотидов с 5'- и 3'- концов молекулы. В некоторых случаях из одной длинной мол-лы-предшественника в результате ферментативного расщепления образуются 2 и даже большее число разных тРНК.
Процессинг предшественников мРНК у эукариот – сложный процесс. Эукариотические м-РНК, обнаруживаемые в цитоплазме, обладают тремя отличительными свойствами: 1) Эукариот мРНК являются моногенными мол-ми, в то время как многие прокариотич мРНК – полигенны 2) Эукариот мРНК содержат в своём 3'-конце «хвост» из 100-200 последоват присоединён остатков А, так называемый poly-А-хвост. Этот хвост синтезируется отдельно из мол-л АТР с помощью полиденилат-полимеразы. 3) Отличительной особенностью большинства эукариот мРНК – это наличие в них 5'-концевого «кэпа», представляющего собой остаток 7-метилгуанозина. «кэп» возможно принимает участие в связывании м-РНК с рибосом, инициируя процесс трансляции. Не исключ также, что «кеп» и poly(А)-хвост предохраняют м-РНК от ферментат разрушения. В эукариот ядре мРНК должны пройти дальнейшие стадии процессинга, заключающиеся в удалении интронов. В ядре присутств РНК особого класса, которая при синтезе белка очень быстро обновляется. Эта РНК носит название гетерогенной ядерной РНК (гяРНК) и состоит из смеси очень длинных мол-л РНК. гяРНК служат предшественниками цитоплазматических м-РНК. гяРНК по размеру гораздо больше, чем зрелые мРНК, к тому же для первой характерно большее разнообразие нуклеотидных последовательностей, чем для второй. Процессинг предшественников мРНК. Роль мяРНК в вырезании интронов и воссоединении экзонов. Довольно долго эта проблема казалась загадочной, но её решение стало реальным после открытия в эукариот генах нетранслируемых вставочных последовательностей, или интронов. После обнаружения в ДНК интронов естественно встал вопрос: транскрибируются ли они коллинеарным образом вместе с экзонами, образуя очень длинный предшеств мРНК, комплимент и коллинеарн как экзонам, так и интронам, или же РНК-полимераза «перетаскивает» через интроны и транскрибирует только экзоны. Оказалось, что эукариотическая РНК- полимераза транскрибирует и экзоны и интроны, причем точно в той последовательности, в которой они находятся в гене. При этом образуются очень длинные предшественники РНК, которые содержат участки нетранслируемой РНК, комплементарные нуклеотидным последовательностям интронов ДНК. Такие предшественники мРНК представляют собой значительную часть гяРНК и обнаруживаютя только в ядре. Кроме того, именно наличием интронов объясняются структурные соотношения между гяРНК и соответствующими цитоплазматическими мРНК. Удаление нетранслируемых интронов при процессинге предшественников иРНК протекает таким образом, что следующие друг за другом экзоны, т.е. кодирующие фрагменты мРНК, иногда физически не разобщаются. Экзоны очень точно соединяются между собой с помощью мол-л другого класса РНК, присутствующих в ядре и называемых малыми ядерными РНК (мя РНК). Функцию этих коротких ядерных РНК, состоящих приблизительно из ста нуклеотидов, удалось установить после того, как было обнаружено, что их нуклеотидная последовательность комплиментарна последовательностям на концах каждого из интронов. В результате спаривания оснований, содержащихся в мяРНК и на концах свернутого в петлю интрона, последовательности двух экзонов сближаются т.о., что становится возможным удаление разделяющего их интрона и ферментативное соединение (сплайсинг) кодирующих ферментов (экзонов). Т.о., мол-лы мяРНК играют роль временных матриц, удерживающих близко друг от друга концы двух экзонов для того, чтобы сплайсинг произошел в правильном месте.
Роль мяРНК в вырезании интронов и воссоединении экзонов. После того как таким путём из РНК удаляются все интроны и тем самым завершается процессинг предшественника мРНК, зрелая мРнк покидает ядро. Чтобы сделать это, мРНК сначала связывается с двумя специальными белками, которые проводят мРНК в цитоплазму сквозь поры в ядерной оболочке. Эти поры, окружены сложным ансаблем белковых молекул, пропускают из ядра, по-видимому, только полностью «созревшие» мРНК. Обрывки РНК, оставшиеся после процессинга, расщепляются нуклеазами. Образовавшиеся при этом нуклеозид-5'-монофосфаты переводятся с помощью АТР в нуклеозид-5'-трифосфаты и вновь используются для синтеза РНК в ядре. В предшественниках тРНК наряду с удалением концевых последовательностей происходят изменения двоякого рода. Во-первых: к некоторым тРНК присоединяется 3'-концевая тринуклеотидная последовательность –С-С-А (3'); в других тРНК этот 3'-концевой тринуклеотид уже содержится в транскрипте. 3'-концевой остаток А представляет собой именно ту часть мол-лы тРНК, с которой ковалентно связывается соответствующая ей аминокислота перед включением в растущую полипептидную цепь на рибосоме. Во-вторых: ряд оснований в тРНК специфическим образом модифицируется одни метилируются, другие дезаминируются, третьи восстанавливаются.
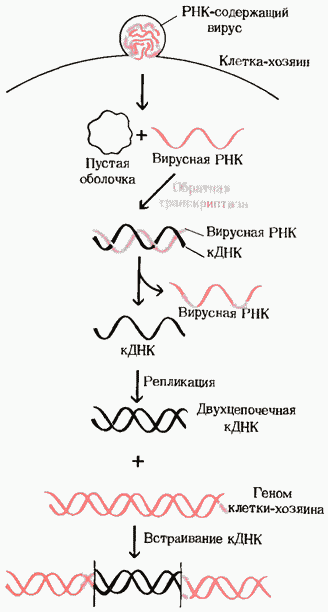
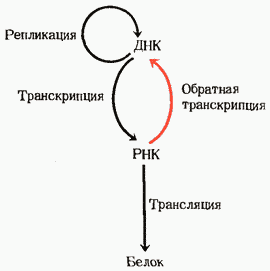
19. Обратная транскрипция. Некоторые онкогенные РНК-содержащие вирусы животных, такие, как вирус саркомы Рауса, имеют уникальный фермент - РНК-зависимую ДНК-полимеразу, часто называемую обратной транскриптазой. После того как такой вирус попадает в клетку-хозяина, этот фермент способен катализировать синтез ДНК, комплементарной по отношению к вирусной РНК, которая играет при этом роль матрицы. В рез-те образ-ся ДНК, к-ая содержит гены, обусловливающие рак; эта ДНК часто встраивается в геном эукариотической клетки-хозяина, где она может в течение многих поколений оставаться в скрытом, т. е. неэкспрессируемом, состоянии. При определенных условиях такие бездействующие вирусные гены могут активироваться и вызывать репликацию вируса; при других же условиях они могут способствовать превращению такой клетки в раковую. Существование обратных транскриптаз в РНК-содержащих онкогенных вирусах было предсказано еще в 1962 г. Говардом Темином из Висконсинского университета, а наличие их в этих вирусах было в конце концов продемонстрировано в 1970 г. как самим Темином, так и независимо от него Дэвидом Балтимором из Массачусетского технологического института. Их открытие привлекло большое внимание главным образом потому, что оно представляло собой доказательство возможности передачи генетической информации в направлении от РНК к ДНК. Оно позволило представить, каким способом включаются в геном клетки-хозяина онкогены, находящиеся в РНК-содержащих вирусах в виде РНК. Именно благодаря этому открытию пришлось по-иному сформулировать центральную догму молекулярной биологии. Обладающие обратной транскриптазой РНК-содержащие вирусы называют также ретровирусами («ретро» по-латыни означает «назад»).
Рис. Участие обратной транскриптазы в образовании комплементарной ДНК на вирусной одноцепочечной РНК-матрице в животной клетке. Полученная кДНК может встраиваться в геном клетки-хозяина.
Рис. Расширенное толкование центральной догмы молекулярной генетики, учитывающее возможность передачи генетической информации от РНК к ДНК в связи с открытием обратных транскриптаз.
|