Деление клеток эукариот
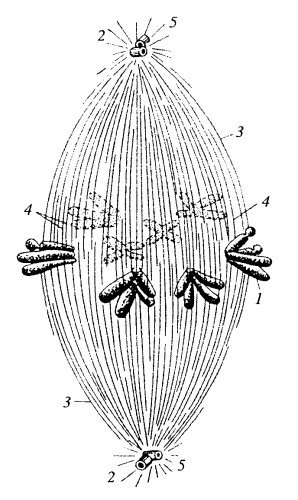
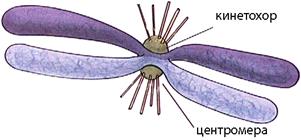
В случае эукариотической клетки, которая содержит множество разнообразных органелл, возникает проблема распределения между дочерними клетками различных клеточных структур. Компоненты цитоскелета и одномембранные структуры могут разбираться на фрагменты и собираться заново, поэтому нет проблемы передачи этих структур дочерним клеткам: они возникают заново. Митохондрии и пластиды содержат свой собственный геном, поэтому могут образовываться путем деления предсуществующих митохондрий и пластид. Но они делятся независимо от деления ядра клетки, и количество таких органелл в клетке достаточно велико (клетка печени содержит более 1000 митохондрий), поэтому при случайном распределении они попадают в обе дочерние клетки. Проблема возникает при распределении наследственного вещества. Оно представлено у эукариот несколькими или многими линейными молекулами, каждая их которых образует хромосому. Каждая дочерняя клетка при делении должна получить одну полную копию генетического материала, представленного всеми хромосомами. Если какая-то из дочерних клеток после деления получит не весь набор хромосом или лишние хромосомы, то это означает, что в ней произошла геномная мутация — изменение генетического материала. Это может иметь весьма серьезные последствия для клетки, в том числе ее гибель или превращение в раковую клетку. Поэтому в ходе эволюции в клетках эукариот сформировался высокоточный механизм расхождения линейных хромосом к разным полюсам клетки при помощи микротрубочек — белковых нитей цитоскелета. Микротрубочки при делении клетки образуют своеобразную структуру, называемую веретеном деления. Полюса веретена деления представлены клеточными центрами, от которых расходятся микротрубочки. Клеточные центры играют роль центров организации микротрубочек. У животных и части одноклеточных в каждом из клеточных центров находится по две центриоли (это структуры из микротрубочек, см. тему «Немембранные органеллы»). Часть микротрубочек, отходящих от каждого из полюсов, встречается и контактирует посередине клетки. Они участвуют в расталкивании полюсов друг от друга, чтобы они вместе с отошедшими к ним хромосомами в результате деления попали в разные клетки. Часть микротрубочек присоединяется к первичным перетяжкам, или центромерам, хромосом. Каждая хромосома имеет в области центромеры специфический белковый комплекс для прикрепления микротрубочек — кинетохор.
Рис. 2: 1 — хромосомы; 2 — клеточный центр (2 центриоли); 3–5 — микротрубочки веретена
Рис. 3 Наличие веретена деления является отличительной особенностью деления эукариотических клеток по сравнению с прокариотами. Центриоли удваиваются между делениями клетки. После деления в клетке есть 1 клеточный центр, содержащий 2 центриоли. Затем центриоли немного расходятся, и около каждой из них начинается формирование перпендикулярной дочерней центриоли. В результате перед делением в клетке оказывается 2 пары центриолей (т. е. всего 4) в виде двух клеточных центров, которые расходятся и становятся полюсами веретена деления. Существует два типа деления клеток эукариот: митоз и мейоз. При митозе хромосомный набор (плоидность) клетки не меняется, обе дочерние клетки полностью генетически идентичны исходной. Это обычный способ деления клеток, например, при формировании тел многоклеточных животных и растений. В результате мейоза, который включает в себя 2 деления, из одной диплоидной клетки получается 4 гаплоидных, причем все они генетически отличаются друг от друга. Это может происходить при формировании гамет или спор. В некоторых книгах можно встретить упоминание третьего типа деления эукариот — амитоза, под которым подразумевается прямое деление ядра клетки без участия микротрубочек и веретена деления. В настоящее время все явления, которые описывались ранее как амитоз, считаются либо артефактами (то есть следствиями повреждения клеток при приготовлении препаратов), либо следствиями апоптоза (запрограммированной клеточной смерти) и других известных процессов. В неделящейся клетке хромосомы не различимы, хотя они присутствуют в ядре. Так происходит потому, что плотность упаковки генетического материала в клетке может быть разной. Максимально плотными хромосомы становятся в начале деления. Поэтому при делении хромосомы в клетке различимы даже в световой микроскоп. После деления упаковка хромосом становится менее плотной, поэтому мы перестаем видеть их границы, хотя они сохраняют свою целостность, но находятся в состоянии «размотанного клубка». Однако их все-таки можно увидеть при помощи специфических методов визуализации хромосом. Совокупность ДНК и связанных с ней белков в неделящейся клетке, которая представляет собой деспирализованные хромосомы, называется хроматином. Почему так происходит? Дело в том, что в том состоянии, в котором хромосомы находятся при делении, генетический материал упакован настолько плотно, что он не может работать. Не может происходить транскрипция, репликация. Да это и не нужно. Главная задача при делении — быстро распределить хромосомы по дочерним клеткам. После деления хромосомы уже не могут оставаться в таком плотно упакованном состоянии, иначе клетка не сможет ими «пользоваться», и ее жизнедеятельность будет невозможна. Поэтому хромосомы «распаковываются», декомпактизуются. Можно сравнить эти процессы с упаковкой вещей при переезде и их распаковкой после переезда в новую квартиру. В начале деления происходит, таким образом, конденсация хроматина, или спирализация хромосом (это одно и то же), а в конце деления — деконденсация хроматина, или деспирализация хромосом.
|