Введение.
Тема стволовых клеток и их использование в регенеративной медицине для восстановления формы и функции повреждённых тканей достаточно хорошо раскрыта. Сейчас речь пойдёт о нише стволовых клеток, т.е. о тех "обычных" соматических клетках, которые окружают пул стволовых клеток, непосредственно взимодействуют с ними и тем самым регулируют пролиферативный потенциал стволовых клеток путём межклеточного взаимодействия.
Концепция ниш стволовых клеток
В 1978 году R. Sсhofield опубликовал статью в журнале "Blood cells", в которой он теоретически обсуждает роль клеток окружения в регуляции жизнедеятельности стволовых клеток. Примерный перевод основного тезиса этой статьи звучит так:
"здесь предлагается гипотеза, по которой необходимо рассматривать стволовые клетки вкупе с окружающими их клетками, которые непосредственно влияют на их функционирование. Это своего рода "тканевая" единица и должна рассматриваться целостно с учётом окружающих соматических клеток. Бесперебойная работа этой единицы обеспечивает нормальное деление и дифференцировку стволовых клеток внутри неё. Однако, как любая ткань, и эта единица подвержена старению и потенциал деления и функциональные возможности окружающих клеток является лимитирующим и определяющим судьбу стволовых клеток внутри неё."
Понятие "ниши" было введено позже, термин был взят из учения об экологии и, в настоящее время, определяется как "ограниченное специализированное микро-окружение, которое интегрирует и осуществляет межклеточные сигналы для регуляции и поддержании гомеостаза принадлежащим ей стволовым клеткам".
 Рисунок 1.
Рисунок 1.
Стволовые клетки не существуют в организме сами по себе, они находятся в определенном микроокружении, которое обычно обозначают термином ниша.
В настоящее время этот термин также используется для обозначения совокупности факторов, обеспечивающих жизнеспособность и самовоспроизведение стволовых клеток и дифференциацию дочерних транзиторных клеток. Среди этих факторов следует упомянуть наличие базальной мембраны, молекул внеклеточного матрикса и присутствие соседних клеток, продуцирующих факторы роста и другие регуляторные молекулы.
Ниша активно участвует в регуляции пролиферации и дифференцировке стволовых клеток, она обеспечивает самоподдержание стволовых клеток и длительное их пребывание в состоянии покоя. Стволовые клетки прочно закреплены в нише молекулами адгезии, в частности интегринами(это трансмембранные гетеродимерные клеточные рецепторы, взаимодействующие с внеклеточным матриксом и передающие различные межклеточные сигналы. От них зависит форма клетки, её подвижность, они участвуют в регулировкеклеточного цикла.)
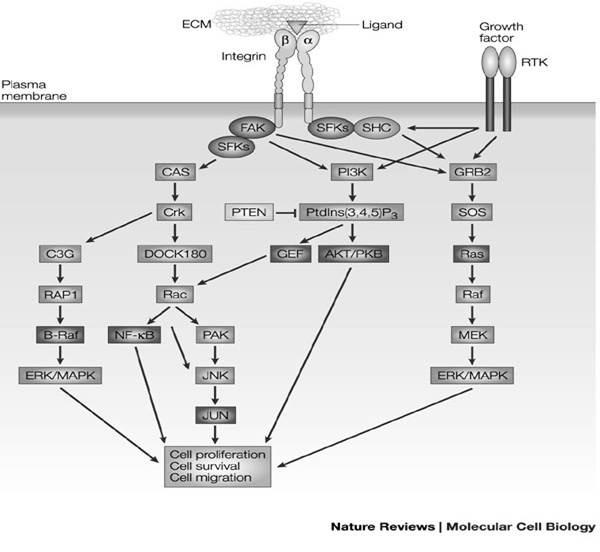 Рисунок 2.
Рисунок 2.
В то же время свободные стволовые клетки могут находить путь в соответствующую нишу благодаря хемотаксису(направленное движение фагоцита по отношению к объекту фагоцитоза). Ниши являются частью структурно-функциональных единиц, из которых состоят ткани.
Ниша стволовых клеток может оставаться свободной и в дальнейшем ее могут занять новые клетки. Пустые ниши могут существовать независимо от стволовых клеток и при трансплантации в них стволовых клеток обеспечивать их нормальное функционирование.
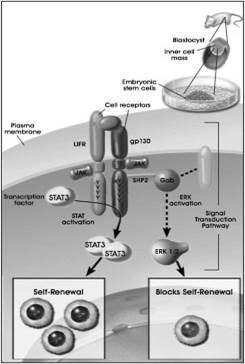 Рисунок 3.
Рисунок 3.
Одно из назначений ниши в тканях взрослого организма заключается в ограничении пролиферации стволовых клеток только необходимостью поддерживать тканевой гомеостаз. Другое назначение ниши - создание условий для максимальной защищенности стволовых клеток от внешних воздействий.
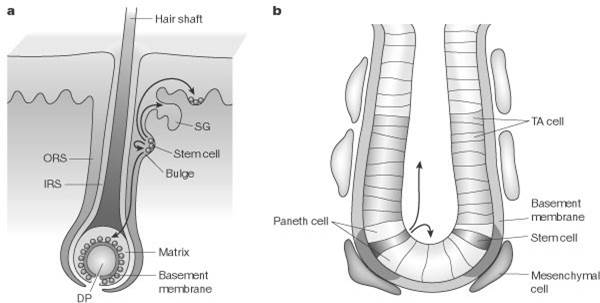
Например, стволовые клетки эпителия кишечника находятся в нижней части крипт, в волосяном фолликуле стволовые клетки локализуются под
сальной железой, стволовые клетки роговицы - в области лимба.
Переход стволовых клеток в состояние покоя также повышает их устойчивость к внешним воздействиям.
Для самоподдержания стволовые клетки должны получать от своего
микроокружения (ниши) определенные сигналы. Например, для поддержания эмбриональных стволовых клеток мыши в недифференцированном состоянии in vitro необходимфактор ингибирования лейкемии (LIF).
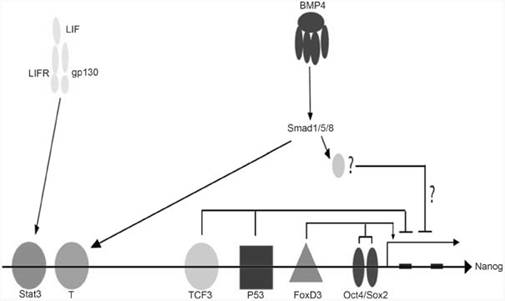
Внутриклеточные сигналы также необходимы для поддержания неограниченной пролиферации стволовых клеток. Для поддержания плюрипотентности и предотвращения дифференцировки эмбриональных стволовых клеток необходима экспрессия транскрипционного фактора Nanog.(это транскрипционный фактор, участвующий в самообновлении недифференцированных эмбриональных стволовых клеток.)
Рисунок 4.
В результате деления стволовые клетки дают начало дочерним клеткам с коротким клеточным циклом. Дочерние клетки благодаря нескольким
последовательным делениям создают большой компартмент (популяцию) транзиторных клеток, которые затем превращаются в дифференцированные клетки, выполняющие специфические функции в организме.
Рисунок 5.
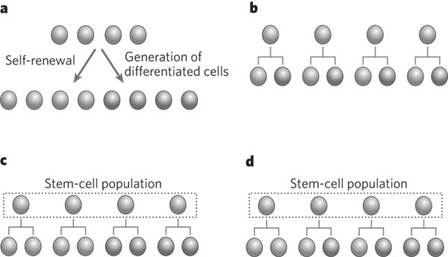
Образование большого количества дифференцированных клеток обеспечивается именно за счет размножения транзиторных клеток при малом числе делений стволовых клеток.
Это позволяет уменьшить риск генетических нарушений, которые могут
произойти в процессе репликации и пролиферации стволовых клеток, поскольку именно с генетическими нарушениями стволовых клеток связан неопластический рост. Генетические нарушения в транзиторных клетках представляют меньшую опасность, поскольку эти клетки, как правило, прекращают пролиферацию и дифференцируются.
Длительное время существовало убеждение, что транзиторные клетки необратимо выходят из компартмента стволовых клеток, однако постепенно стали появляться данные, свидетельствующие о том, что между стволовыми и транзиторными клетками нет резкой границы, а скорее имеется постепенный переход. После того как стволовые клетки в результате деления дают начало транзиторным клеткам, последние еще некоторое время могут сохранять свойства стволовых клеток. Ведущий учёный в этой области, C. Potten, глава биотех-компании Epistem, называет их потенциальными стволовыми клетками и считает, что в норме они относятся к транзиторной популяции, но при определенных условиях, например, гибели пред существующих стволовых клеток, могут заместить последние.
Представляют интерес экспериментальные данные относительно большой пластичности транзиторных клеток. Транзиторные клетки эпителия роговицы находятся в ее центральной части и легко могут быть отделены от стволовых клеток, которые локализованы в области лимба.
Рисунок 6.
Было установлено, что транзиторные клетки эпителия роговицы кролика
могут быть перепрограммированы при взаимодействии с дермой эмбрионов мыши (дорсальной, верхней губы и подошвы). Полученные результаты показывают, что клетки роговицы взрослого животного отвечают на специфические стимулы эмбриональной дермы. Сначала появляется новый базальный слой клеток, в котором не экспрессируются кератины роговицы, а затем появляются пилосебацейные единицы, или потовые железы, в зависимости от типа дермы, и наконец в верхних слоях клеток появляется экспрессия кератинов эпидермального типа. Таким образом, происходит перепрограммирование клеток роговицы.




