Дробление
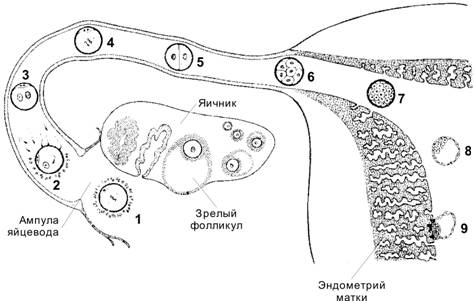
Дробление — митотическое деление диплоидных клеток без увеличения их суммарного объёма — стадия развития, наступающая после оплодотворения. При дроблении происходит относительно быстрое увеличение количества клеток (бластомеры), от 2 до примерно 12–16 к третьим суткам после оплодотворения, когда концептус достигает стадии морулы и выходит в полость матки из маточной трубы (рис. 3-6). Сигнальный белок дробления. В зиготе обнаружены два принесённые сперматозоидом белка с Mr 14 и 18 кД, содержащие одну и ту же Аг-детерминанту. АТ к этому Аг блокируют первые дробления зиготы, не оказывая влияния на другие процессы. Дробление отличается от обычного клеточного цикла отсутствием фаз G1 и G2 и очень короткой S–фазой, во время которой происходит синтез ДНК. Интервал между делениями дробления составляет примерно 12–24 часа. Роста клеток не происходит и в ходе дроблений клетки уменьшаются в размерах, так что диаметр концептуса не превышает 100 мкм. На стадии бластоцисты дробления прекращаются и клетки возобновляют нормальный клеточный цикл G1, S, G2, M. В ходе дробления и на последующих стадиях развития происходит устранение дефектных клеток путём апоптоза. На всех стадиях развития наблюдается конститутивная экспрессия белка BAX. В ходе развития частота экспрессии проапоптозных и антиапоптозных белков семейства BCL-2 проявляется сходным образом, но на 2-клеточной стадии преобладает экспрессия проапоптозных белков. В отличие от нормальных, клетки дефектных концептусов на стадии 4-х бластомеров экспрессируют BAK. Характер дробления определяется количеством и распределением в цитоплазме желточных включений. У человека — плацентарного млекопитающего — в связи с внутриутробным питанием плода через плаценту нет необходимости в создании больших запасов желтка. Поэтому желток распределён в цитоплазме равномерно (изолецитальная яйцеклетка). Зигота, образованная такой яйцеклеткой, совершает голобластическое дробление, т.е. полностью разделяется на два бластомера. Дальнейшие дробления — асинхронные и несколько неравномерные. При дроблении зигота окружена прозрачной оболочкой, сохраняющейся до стадии бластоцисты.
Рис. 3-6. Концептус от оплодотворения до имплантации. Овуляция наступает на 14-й день менструального цикла, в течение 1 сут происходит оплодотворение. Трое суток с момента оплодотворения концептус продвигается по маточной трубе, на 4 сут попадает в полость матки и через 5,5–6 сут имплантируется в эндометрий. В ходе перемещения по маточной трубе происходит дробление, и в полости матки оказывается бластоциста, готовая к имплантации. 1 — овоцит тотчас после овуляции, 2 — оплодотворение через 12–24 ч после овуляции, 3 — стадия появления мужского и женского пронуклеусов, 4 — начало дробления, 5 — двухклеточная стадия (30 часов), 6 — морула, состоящая из 12–16 бластомеров (3 сут), 7 — продвинутая морула в полости матки (4 сут), 8 — ранняя бластоциста (4,5–5 сут), 9 — имплантация бластоцисты (5,5–6 сут). [17]
Рис. 3-7. Компактизация и морула на ранних этапах развития. В результате компактизации уменьшается размер межклеточных пространств между бластомерами, они сближаются, и формируется морула. В моруле различают внутреннюю часть (клетки связаны щелевыми контактами) и наружную часть (клетки соединены при помощи плотных контактов). Из клеток внутренней части морулы развивается эмбриобласт, а из клеток наружной части формируется трофобласт. Внутренняя клеточная масса и трофобласт — главные структурные элементы бластоцисты. [17] Первое дробление (рис. 3-6) начинается примерно через 30 часов после оплодотворения. Плоскость первого деления проходит через область направительных телец. Поскольку желток в зиготе распределён равномерно, выделение анимального и вегетативного полюсов затруднено. Область отделения направительных телец обычно рассматривают как анимальный полюс. Образующиеся два бластомера несколько различны по размерам. В результате первого деления дробления нарушается симметрия зародыша и образуется два бластомера с различными потенциями развития: один бластомер дает начало «эмбриональной части» бластоцисты (полярный трофобласт и глубокая часть внутренней клеточной массы), а другой бластомер — «неэмбриональной части» бластоцисты (муральный трофобласт и поверхностный слои внутренней клеточной массы). Второе дробление (рис. 3-6). Образование второго митотического веретена в каждом из бластомеров происходит вскоре после окончания первого деления, плоскость второго деления проходит перпендикулярно плоскости первого деления дробления, концептус переходит в стадию 4-х бластомеров. Однако, дробление у человека асинхронно, поэтому при переходе от стадии 2-х бластомеров к стадии 4-х бластомеров в течение некоторого времени можно наблюдать трёхклеточный концептус. На стадии 4-х бластомеров синтезируются все основные типы РНК. Третье дробление. На этой стадии несинхронность дробления проявляется в большей мере, в итоге образуется концептус с различным количеством бластомеров; условно его можно отнести к стадии 8 бластомеров (рис. 3-6, рис. 3-7). До этого бластомеры расположены рыхло. Непосредственно перед следующим (4-м) делением дробления с образованием 16 бластомеров эмбрион подвергается компактизации (рис. 3-8). Концептус уплотняется (компактизация), поверхность соприкосновения бластомеров увеличивается, объём межклеточного пространства уменьшается. Сближение бластомеров в ходе компактизации — необходимое условие для образования между бластомерами специализированных межклеточных контактов (щелевых и плотных), формирующихся на поздней 8-клеточной стадии (морула). До формирования контактов в плазматическую мембрану бластомеров начинает встраиваться E-кадгерин (увоморулин) — белок адгезии клеток. В бластомерах ранних концептусов E-кадгерин равномерно распределён в клеточной мембране. Позднее в области межклеточных контактов образуются скопления (кластеры) молекул E-кадгерина.
Рис. 3-8. Молекула E-кадгерина в бластомере связана с примембранным цитоскелетом (актином) через катенин. Клетки начинают экспрессировать Ca2+-зависимый белок адгезии E-кадгерин, который способствует сближению (компактизации) и поляризации клеток. [102] В ходе компактизации E-кадгерин способствует поляризации клеток. Между апикально-латеральными областями наружных бластомеров формируются плотные контакты, достигающие максимального развития на 32-клеточной стадии, когда начинает формироваться полость (бластоцель). Позднее, в ходе гаструляции, E-кадгерин замещаетсяN–кадгерином. МОРУЛА По мере увеличения числа бластомеров концептус перемещается по маточной трубе и к третьим суткам достигает стадии морулы (рис. 3-6). Морула — группа клеток, возникших в ходе нескольких делений дробления и заключённых внутри прозрачной оболочки (рис. 3-7). Центрально расположенные клетки морулы образуют щелевые контакты, при помощи которых осуществляются информационные межклеточные взаимодействия. Именно из этих клеток и возникает собственно эмбрион. Периферические клетки морулы соединяют плотные контакты; они формируют барьер, обособляющий внутреннюю среду морулы. Для внутренних клеток морулы характерна выраженная экспрессия Nanog. LIF/STAT3-независимый фактор транскрипции Nanog необходим для самоподдержания эмбриональных стволовых клеток и сохранения их плюрипотентности. В ходе дальнейшего развития nanog (ген плюрипотентности) экспрессируется клетками внутренней клеточной массы бластоцисты. По мере развития экспрессия nanog снижается и сохраняется только в первичных половых клетках. В обособлении в моруле клеток будущей внутренней клеточной массы также участвует фактор транскрипции Oct4, при этом происходит подавление экспрессии генов, ответственных за дифференцировку клеток в трофобласт.Oct4, как и Nanog, отвечает за поддержание плюрипотентности эмбриональных стволовых клеток и в дальнейшемOct4 также экспрессируется клетками внутренней клеточной массы. С другой стороны, на стадии поздней морулы поляризованные бластомеры наружного слоя экспрессируют cdx2(c au d al-related homeobo x gene). Повышенная экспрессия фактора транскрипции Cdx 2 индуцирует формирование трофобласта. Гены Cdx1 и Cdx2, кодирующие соответствующие факторы транскрипции, гомологичны гену, содержащему гомеобокс, Caudal у Drosophila. Cdx 1 экспрессируется на ранних стадиях развития, когда определяются оси тела; на более поздних стадиях экспрессия Cdx 1 характерна только для клеток эмбриональной энтодермы, дающих начало кишечной трубке. У взрослого человека Cdx 1 экспрессируется стволовыми эпителиальными клетками крипт кишечника.
|