TABLE 3 Dextran-binding receptors: roles in infections
DC, dendritic cell; DC-SIGN, dendritic cell–specific intercellular adhesion molecule (ICAM)-3-grabbing nonintegrin; IFN, interferon; L-SIGN, liver/lymph node-specific intercellular adhesion molecule (ICAM)-3-grabbing nonintegrin; SARS, severe acute respiratory syndrome; SIV, simian immunodeficiency virus.
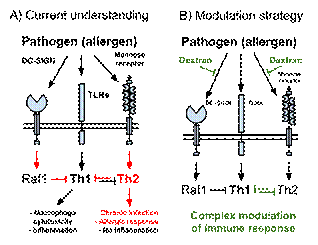
Dextran-isoniazid has shown interesting results in a model of tuberculosis-like granulomatosis induced by Bacillus Calmette–Guérin (BCG) injection. The intensity of fibrotic lesions in this model after treatment with dextran conjugate was compared with free isoniazid treatment. Fibrosis of the lung decreased 30%, of the spleen 3.5-fold, and of the liver more than fourfold. Hepatotoxicity decreased 2.2-fold, and the development of necrosis into granulomas decreased 10-fold (159). Decreased lung remodeling may be beneficial for prevention of caviation and subsequent transmission (160) of tuberculosis, and could also help drugs reach the mycobacteria inside granulomas, that is itself an important problem (161). Dextran influences the phagosomal-lysosomal fusion and the death rate of mycobacteria BCG inside mouse peritoneal macrophages. The control rate of death inside macrophages was 33%, and with dextran (22 mg/ml) it was 39%. Isoniazid treatment (7 mg/ml) yielded a bacterial death rate of 43%, while the conjugate of dextran with isoniazid (25 mg/ml, same isoniazid content) yielded a 53% death rate. The latter result may be explained by targeted delivery of dextran into the phagosomes and lysosomes where the pathogen is taken up (162). An increase in phagocytic activity after dextran uptake is probably connected with NADPH oxidase 2 upregulation which is responsible for antimicrobial activity (163). In the systemic candidiasis model the dextran-amphotericin B conjugate given 10 days after infection decreased the number of granulomas in the liver by fourfold (164). In experiments on dextran–rimantadine this conjugate has shown to have a significantly better defencive effect in the chicken embryo and mouse models for influenza A and B virus and in the mouse model of tick-borne encephalitis (165). It remained unclear whether dextran alone could cause similar effects in the treatment of infections. Regularly infused in mice in a model of BCG-induced granulomatosis, oxidized dextran (OD; in these studies-the molecule of clinical dextran containing less than 3% of glucose units oxidized with formation of aldehyde groups) reduced the number and size of granulomas in the organs; increased numbers of fibroblasts (with reduced activity) in the granulomas; decreased destructive and necrotic changes in the liver; and decreased fibrosis in the liver and lungs (166). In a mouse influenza model, OD decreased fatality by 3.3-fold and significantly decreased lung fibrosis (167). In a model of systemic candidiasis, the number of granulomas in the brain decreased eightfold after OD treatment compared with antifungal amphotericin B. While the control group of mice died, 60% of OD-treated mice survived (168). The mechanism of OD action is still undiscovered; however, this form of dextran has been shown to increase the degree of adhesion of peritoneal cells, which may indicate increased activity of macrophages (169). OD reduces the viability of these cells, but conversely it stimulates metabolic and oxidative processes (169). In vitro dextran, and to a greater extent OD, are able to stimulate macrophage production of granulocyte-macrophage colony-stimulating factor (169), which supports the differentiation and activation of antigen-presenting cells (170). OD causes a shift in the balance of activities between nitric oxide synthase and arginase towards increasing nitric oxide production by macrophages (171). Another effect is increased macrophage ROS production (172). Chemical differences between dextran and OD are not significant; it is unknown whether oxidation played a role in in vivo results. Probably specific binding of MR and DFRs by dextran modulates pathogen-induced T helper responses (Figure 4) (173, 174). Thus antifibrotic action of dextran in BCG model (159, 166) could be linked to restricted Th2 reaction contributing to tissue remodelling. If this hypothesis is true, dextran could also modulate the immune response to Th2 overreaction-inducing allergens dependent on MR (175) and DC-SIGN (176, 177). Preliminary results are available concerning the in vivo action of nonmodified dextran in models of infections dependent on dextran-binding receptors. Dextran introduced intranasally simultaneously with heat-killed M. tuberculosis H37Rv decreased lung concentrations of both IFN-g and IL-10, while the IFN-g/IL-10 ratio decreased 2.5-fold, a result that rather illustrates suppression of Th1 response (178). Dextran introduced intranasally simultaneously or a day before infection with 10 LD50 of the H5N1 influenza virus saved or prolonged lives of mice (179). These experiments do not provide evidence on dextran’s mechanisms of action, a question that will be addressed in future works. They show, however, that dextran may be a promising molecule to add to the long list of treatments against infections dependent on dextran-binding receptors (Table 3).
Figure 4. Dextran and glycan-lectin interactions. This simplified scheme shows that if dextran decrease the availability of MR and DC-SIGN for the pathogens, this may influence immune responses. It is known that DC-SIGN ligands prevent binding and entry of pathogens, interfere with trans-infection of T cells by DCs, skew the myeloid cells activation phenotypes and influence immune response.
DEXTRAN IN PREVENTING HIV INFECTION AND TRANSMISSION Sexual transmission of HIV is the most prevalent route for infection (180, 181). DCs of intestinal and genital mucosae express DC-SIGN (21). They can be productively infected with HIV and have high capacity to trans-infect the T cells―the main HIV targets. DC-SIGN itself is an important player in the formation of DC-T cell infectious synapses (182, 183); signaling via DC-SIGN promotes increased viral uptake (184) and productive infection (185), and also influences DCs regulatory roles (30). HIV entry inhibitors are commonly used antiretrovirals (186), but there are still no inhibitors of HIV-DC-SIGN interaction introduced into the clinics, in spite of proven importance of receptor in myeloid cells infection and trans-infection of T cells. Dextran 60 given before and after infection provides significant decrease of the HIV-1 viral RNA inside the B-THP-1/DC-SIGN cells. Dextran oligomers also inhibit infection (S. Pustylnikov and P. Jain, unpublished results) and indeed carbohydrate-binding domain of DC-SIGN binds to ~3 carbohydrate units (187). This suggests dextran is an effective inhibitor of HIV-DC-SIGN interaction. It was shown that dextran decreases the mortality rate of HIV-infected human monocyte-derived macrophages from 84% to 48% (188). This could be a result of the inhibition of the minor HIV-DC-SIGN binding (189), as well as a result of the inhibition of HIV-MR interaction shown in macrophage infection and viral transmission (98). We suggest that dextran as a DC-SIGN and MR ligand could not only decrease the rates of HIV infection and trans-infection in myeloid cells, but could also serve to deliver the antiretrovirals or vaccines to DCs. Anti-HIV gel formulations have proven their efficiency in clinical trials (190); use of viral entry inhibitors in gel formultions can provide full protection in vivo (191). If dextran proves to be an HIV entry inhibitor, it could be used as a gel formulation.
CONCLUSIONS The combination of dextran properties is unique. Dextran is a hydrophilic, nonionic molecule with adjustable molecular mass distribution (Figure 2) and viscosity/density in solutions. Dextran’s lack (or near lack) of toxic effects, pyrogenic or allergic reactions and accumulation in the body; its thermal and chemical stability allowing sterilization and obtaining the derivatives; its applicability in mass production at comparably low costs (82, 192): all make dextran an appealing biopolymer for multiple applications. Antimicrobial strategies that could exploit dextran is a speculative topic due to the lack of data. Despite lack of direct evidence, dextran's applications as shown in Figure 2 can be numeous in fields of research and medicine where dextran is applied. Dextran is a popular component of conjugates and nano-particles. Numerous works on drug-dextran conjugates show interesting results in vitro and in vivo and provide arguments for improved pharmaceutical properties of such compounds (reviewed in (193-198). Our analysis suggests that concept of targeted delivery―the conjugation of dextran with antimicrobials to reach the pathogens inside the specific cells that take up dextran (liver cells, macrophages and DCs)―being itslef not a new idea, can benefit from knowledge of dextran-binding receptors and their roles in a number of infections. Dextran’s influences on infections has not been studied comprehensively to date and only minor influences are known. Dextran-binding MR, DC-SIGN (in human)/SIGN-R1/SIGN-R3 (in mice), L-SIGN, and langerin play large roles in infectious diseases (Table 3). Besides regulation of immune cell interplay, these receptors participate in binding, recognition, and uptake of different pathogens. Targeting of dextran-binding receptors (e.g., MR and DC-SIGN) is a popular concept. In recent years studies devoted to the development of DC-SIGN therapeutic ligands have yielded new data in cell biology (203), immunology (204), and biochemistry (205, 206). The concept of therapeutic DC-SIGN antagonists/inhibitors is promising and in need of further development (9, 207). Targeting the MR is suggested for vaccine development (201), for delivery of cargo into macrophages (202) or liver cells (195). Dextran can play a role in the prevention of pathogen binding, entry and signaling in MR-expressing myeloid cells wich participate in blood-brain barrier disruption in neuroinvasive infections (208): this was probably the case in prevention of C. albicans infection in the brain (168). Skewing the T helper responses could be a mechanism that allowed dextran derivatives to decrease tissue remodelling in the BCG infection model (159, 166) (Figure 4). Dextran has been recently used as a backbone for the nucleic acids delivery conjugate and our analysis could help in the development of this field (199). We also note that dextran could be of use in the glycosilation of adenoviruses used for gene transfer (200), possibly improving the biocompatibility and providing predictable uptake by certain cell types and receptors. Further, the route of delivery of dextran and its derivatives require to be taken into consideration. Infusion will result in primary uptake in the liver, which is not a target of respiratory or mucosal infections. Dextran-based sprays or gels are an option, but they are not helpful in generalized infections. Clinical dextrans with molecular weights in the range 35,000 to 80,000 cannot reach a systemic infection if given orally, but smaller molecules such as dextran with an average molecular weight of 1,000 probably can. Dextrans with high molecular weights induce active endocytosis, while smaller molecules do not (36). They may not only decrease the amount of available dextran-binding receptors on the cell surface but also prevent endocytosis and following recycling of receptors (shown for both MR (209) and DC-SIGN (210)) and keep the cells’ endocytic capacity at its initial level. Medical and biological applications of dextran can be considered in a new way via the prism of receptor-specific interactions. This can be an instrument to interpret the data on dextran conjugates and derivatives. If antimicrobial properties of dextran can be applied in humans, dextran might become an approved, specific, nontoxic, cheap, and accessible immunomodulatory drug. These qualities are extremely important in the case of deadly infections that affect resource-limited populations. Dextran may possess antimicrobial and antiallergic effects owing to binding to MR, DFRs, and langerin. This review suggests a primary aim for future studies: testing of the ability of dextran to act against a panel of pathogens exploiting dextran-binding receptors to enter the cells and to modulate the immune responses.
ACKNOWLEDGMENTS This work was supported in part by Novosibirsk Tuberculosis Research Institute, Novosibirsk, Russia and Scientific Center for Clinical and Experimental Medicine, Novosibirsk, Russia. We thank Stefan Martin from University of Freiburg, Germany, for help in preparing the manuscript and helpful comments. All authors have no potential conflicts of interest to declare.
REFERENCES 1. Atik M. Dextrans, their use in surgery and medicine: With emphasis on the low molecular weight fractions. Anesthesiology. 1966;27:425-38. 2. Sidebotham RL. Dextrans. Advances in carbohydrate chemistry and biochemistry. 1974;30:371-444. 3. Belder AN. Dextran, Handbook. Amersham Biosciences. 2003. 4. Heinze T, Liebert T, Heublein B, Hornig S, Klemm D. Functional Polymers Based on Dextran. Polysaccharides II: Springer Berlin Heidelberg; 2006. p. 199-291. 5. van Kooyk Y, Geijtenbeek TBH. DC-SIGN: escape mechanism for pathogens. Nature Reviews Immunology. 2003;3(9):697-709. 6. Geijtenbeek TBH, Kwon DS, Torensma R, van Vliet SJ, van Duijnhoven GCF, Middel J, et al. DC-SIGN, a Dendritic Cell-Specific HIV-1-Binding Protein that Enhances trans-Infection of T Cells. Cell. 2000;100(5):587-97. 7. Arrighi J-Fo, Pion M, Wiznerowicz M, Geijtenbeek TB, Garcia E, Abraham S, et al. Lentivirus-mediated RNA interference of DC-SIGN expression inhibits human immunodeficiency virus transmission from dendritic cells to T cells. Journal of virology. 2004;78(20):10848-55. 8. Balzarini J, Van Herrewege Y, Vermeire K, Vanham G, Schols D. Carbohydrate-binding agents efficiently prevent dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin (DC-SIGN)-directed HIV-1 transmission to T lymphocytes. Molecular pharmacology. 2007;71(1):3-11. 9. Tran TH, El Baz R, Cuconati A, Arthos J, Jain P, Khan ZK. A Novel High-Throughput Screening Assay to Identify Inhibitors of HIV-1 gp120 Protein Interaction with DC-SIGN. Journal of antivirals & antiretrovirals. 2011;3:49. 10. Jain P, Manuel SL, Khan ZK, Ahuja J, Quann K, Wigdahl B. DC-SIGN mediates cell-free infection and transmission of human T-cell lymphotropic virus type 1 by dendritic cells. Journal of virology. 2009;83(21):10908-21. 11. Kampani K, Quann K, Ahuja J, Wigdahl B, Khan ZK, Jain P. A novel high throughput quantum dot-based fluorescence assay for quantitation of virus binding and attachment. Journal of virological methods. 2007;141(2):125-32. 12. Geijtenbeek TB, van Kooyk Y. Pathogens target DC-SIGN to influence their fate DC-SIGN functions as a pathogen receptor with broad specificity. Apmis. 2003;111:698-714. 13. Chieppa M, Bianchi G, Doni A, Del Prete A, Sironi M, Laskarin G, et al. Cross-linking of the mannose receptor on monocyte-derived dendritic cells activates an anti-inflammatory immunosuppressive program. J Immunol. 2003;171:4552-60. 14. East L, Isacke CM. The mannose receptor family. Biochim Biophys Acta. 2002;1572:364-86. 15. Martinez-Pomares L. The mannose receptor. Journal of Leukocyte Biology. 2012;92(6):1177-86. 16. Zhang XS, Brondyk W, Lydon JT, Thurberg BL, Piepenhagen PA. Biotherapeutic target or sink: analysis of the macrophage mannose receptor tissue distribution in murine models of lysosomal storage diseases. Journal of Inherited Metabolic Disease. 2011;34(3):795-809. 17. Le Cabec V, Emorine LJ, Toesca I, Cougoule C, Maridonneau-Parini I. The human macrophage mannose receptor is not a professional phagocytic receptor. J Leukoc Biol. 2005;77:934-43. 18. Leitinger B, Hohenester E. Mammalian collagen receptors. Matrix Biol. 2007;26:146-55. 19. Sallusto F, Cella M, Danieli C, Lanzavecchia A. Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compartment: downregulation by cytokines and bacterial products. J Exp Med. 1995;182:389-400. 20. Cheong C, Matos I, Choi J-H, Dandamudi DB, Shrestha E, Longhi MP, et al. Microbial Stimulation Fully Differentiates Monocytes to DC-SIGN/CD209< sup>+</sup> Dendritic Cells for Immune T Cell Areas. Cell. 2010;143(3):416-29. 21. Turville SG, Cameron PU, Handley A, Lin G, Pohlmann S, Doms RW, et al. Diversity of receptors binding HIV on dendritic cell subsets. Nature Immunology. 2002;3(10):975-83. 22. Jameson B, Baribaud F, Pohlmann S, Ghavimi D, Mortari F, Doms RW, et al. Expression of DC-SIGN by dendritic cells of intestinal and genital mucosae in humans and rhesus macaques. Journal of virology. 2002;76(4):1866-75. 23. Rappocciolo G, Piazza P, Fuller CL, Reinhart TA, Watkins SC, Rowe DT, et al. DC-SIGN on B lymphocytes is required for transmission of HIV-1 to T lymphocytes. PLoS pathogens. 2006;2(7):e70. 24. Dominguez-Soto A, Sierra-Filardi E, Puig-Kroger A, Perez-Maceda B, Gomez-Aguado F, Corcuera MT, et al. Dendritic Cell-Specific ICAM-3-Grabbing Nonintegrin Expression on M2-Polarized and Tumor-Associated Macrophages Is Macrophage-CSF Dependent and Enhanced by Tumor-Derived IL-6 and IL-10. The Journal of Immunology. 2011;186(4):2192-200. 25. Soilleux EJ, Morris LS, Leslie G, Chehimi J, Luo Q, Levroney E, et al. Constitutive and induced expression of DC-SIGN on dendritic cell and macrophage subpopulations in situ and in vitro. Journal of Leukocyte Biology. 2002;71(3):445-57. 26. Geijtenbeek TB, Torensma R, van Vliet SJ, van Duijnhoven GC, Adema GJ, van Kooyk Y, et al. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune responses. Cell. 2000;100:575-85. 27. Geijtenbeek TBH, Krooshoop DlJEB, Bleijs DA, van Vliet SJ, van Duijnhoven GCF, Grabovsky V, et al. DC-SIGN-ICAM-2 interaction mediates dendritic cell trafficking. Nature immunology. 2000;1(4):353-7. 28. Garcia-Vallejo JJ, van Liempt E, da Costa Martins P, Beckers C, van het Hof B, Gringhuis SI, et al. DC-SIGN mediates adhesion and rolling of dendritic cells on primary human umbilical vein endothelial cells through Lewis< sup> Y</sup> antigen expressed on ICAM-2. Molecular immunology. 2008;45(8):2359-69. 29. Shreffler WG, Castro RR, Kucuk ZY, Charlop-Powers Z, Grishina G, Yoo S, et al. The major glycoprotein allergen from Arachis hypogaea, Ara h 1, is a ligand of dendritic cell-specific ICAM-grabbing nonintegrin and acts as a Th2 adjuvant in vitro. The Journal of Immunology. 2006;177(6):3677-85. 30. Gringhuis SI, den Dunnen J, Litjens M, van Het Hof B, van Kooyk Y, Geijtenbeek TB. C-type lectin DC-SIGN modulates Toll-like receptor signaling via Raf-1 kinase-dependent acetylation of transcription factor NF-kappaB. Immunity. 2007;26:605-16. 31. Dominguez-Soto A, Aragoneses-Fenoll L, Martin-Gayo E, Martinez-Prats L, Colmenares M, Naranjo-Gomez M, et al. The DC-SIGN-related lectin LSECtin mediates antigen capture and pathogen binding by human myeloid cells. Blood. 2007;109(12):5337-45. 32. Powlesland AS, Ward EM, Sadhu SK, Guo Y, Taylor ME, Drickamer K. Widely divergent biochemical properties of the complete set of mouse DC-SIGN-related proteins. J Biol Chem. 2006;281:20440-9. 33. Garcia-Nieto S, Johal RK, Shakesheff KM, Emara M, Royer PJ, Chau DY, et al. Laminin and fibronectin treatment leads to generation of dendritic cells with superior endocytic capacity. PLoS ONE. 2010;5. 34. Viebig NK, Andrews KT, Kooyk Yv, Lanzer M, Knolle PA. Evaluation of the role of the endocytic receptor L-SIGN for cytoadhesion of Plasmodium falciparum-infected erythrocytes. Parasitology Research. 2005;96(4):247-52. 35. Geijtenbeek TB, Groot PC, Nolte MA, van Vliet SJ, Gangaram-Panday ST, van Duijnhoven GC, et al. Marginal zone macrophages express a murine homologue of DC-SIGN that captures blood-borne antigens in vivo. Blood. 2002;100:2908-16. 36. Takahara K, Yashima Y, Omatsu Y, Yoshida H, Kimura Y, Kang YS, et al. Functional comparison of the mouse DC-SIGN, SIGNR1, SIGNR3 and Langerin, C-type lectins. Int Immunol. 2004;16:819-29. 37. Valladeau J, Ravel O, Dezutter-Dambuyant C, Moore K, Kleijmeer M, Liu Y, et al. Langerin, a Novel C-Type Lectin Specific to Langerhans Cells, Is an Endocytic Receptor that Induces the Formation of Birbeck Granules. Immunity. 2000;12(1):71-81. 38. Pipirou Z, Powlesland AS, Steffen I, Pohlmann S, Taylor ME, Drickamer K. Mouse LSECtin as a model for a human Ebola virus receptor. Glycobiology. 2011;21(6):806-12. 39. Mowry RW, Millican RC. A histochemical study of the distribution and fate of dextran in tissues of the mouse. Am J Pathol. 1953;29:523-45. 40. Knolle PA, Germann T, Treichel U, Uhrig A, Schmitt E, Hegenbarth S, et al. Endotoxin Down-Regulates T Cell Activation by Antigen-Presenting Liver Sinusoidal Endothelial Cells. The Journal of Immunology. 1999;162(3):1401-7. 41. Shu SA, Lian ZX, Chuang YH, Yang GX, Moritoki Y, Comstock SS, et al. The role of CD11c+ hepatic dendritic cells in the induction of innate immune responses. Clinical and Experimental Immunology. 2007;149(2):335-43. 42. Grayson MH, Chaplin DD, Karl IE, Hotchkiss RS. Confocal fluorescent intravital microscopy of the murine spleen. Journal of Immunological Methods. 2001;256(1–2):55-63. 43. Mwandumba HC, Russell DG, Nyirenda MH, Anderson J, White SA, Molyneux ME, et al. Mycobacterium tuberculosis Resides in Nonacidified Vacuoles in Endocytically Competent Alveolar Macrophages from Patients with Tuberculosis and HIV Infection. The Journal of Immunology. 2004;172(7):4592-8. 44. Caulfield JP, Farquhar MG. The Permeability of Glomerular Capillaries to Graded Dextrans. Identification of the Basement Membrane as the Primary Filtration Barrier. The Journal of Cell Biology. 1974;63(3):883-903. 45. Broderick C, Duncan L, Taylor N, Dick AD. IFN-Оі and LPS-Mediated IL-10-Dependent Suppression of Retinal Microglial Activation. Investigative Ophthalmology & Visual Science. 2000;41(9):2613-22. 46. Groger M, Holnthoner W, Maurer D, Lechleitner S, Wolff K, Mayr BB, et al. Dermal microvascular endothelial cells express the 180-kDa macrophage mannose receptor in situ and in vitro. The Journal of Immunology. 2000;165. 47. Takahashi K, Donovan MJ, Rogers RA, Ezekowitz RA. Distribution of murine mannose receptor expression from early embryogenesis through to adulthood. Cell Tissue Res. 1998;292:311-23. 48. Bashirova AA, Geijtenbeek TB, van Duijnhoven GC, van Vliet SJ, Eilering JB, Martin MP, et al. A dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin (DC-SIGN)-related protein is highly expressed on human liver sinusoidal endothelial cells and promotes HIV-1 infection. J Exp Med. 2001;193:671-8. 49. Henze E, Schelbert HR, Collins JD, Najafi A, Barrio JR, Bennett LR. Lymphoscintigraphy with Tc-99m-labeled dextran. Journal of nuclear medicine: official publication, Society of Nuclear Medicine. 1982;23(10):923-9. 50. Padera TP, Stoll BR, So PT, Jain RK. Conventional and high-speed intravital multiphoton laser scanning microscopy of microvasculature, lymphatics, and leukocyte-endothelial interactions. Mol Imaging. 2002;1(1):9-15. 51. Ribera J, Pauta M, Melgar-Lesmes P, Tugues S, Fernández-Varo G, Held KF, et al. Increased nitric oxide production in lymphatic endothelial cells causes impairment of lymphatic drainage in cirrhotic rats. Gut. 2013;62(1):138-45. 52. Linehan S. The mannose receptor is expressed by subsets of APC in non-lymphoid organs. BMC immunology. 2005;6. 53. Kato M, Neil TK, Fearnley DB, McLellan AD, Vuckovic S, Hart DN. Expression of multilectin receptors and comparative FITC-dextran uptake by human dendritic cells. Int Immunol. 2000;12:1511-9. 54. Sadaka C, Marloie-Provost M-A, Soumelis V, Benaroch P. Developmental regulation of MHC II expression and transport in human plasmacytoid-derived dendritic cells. Blood. 2009;113(10):2127-35. 55. Xu H, Liew LN, Kuo IC, Huang CH, Goh DL-M, Chua KY. The modulatory effects of lipopolysaccharide-stimulated B cells on differential T-cell polarization. Immunology. 2008;125(2):218-28. 56. Cambi A, Beeren I, Joosten B, Fransen JA, Figdor CG. The C type lectin DC SIGN internalizes soluble antigens and HIV 1 virions via a clathrin dependent mechanism. European journal of immunology. 2009;39:1923-8. 57. Navarrete A-M, Delignat S, Teillaud J-L, Kaveri SV, Lacroix-Desmazes SВb, Bayry J. CD4+ CD25+ regulatory T cell-mediated changes in the expression of endocytic receptors and endocytosis process of human dendritic cells. Vaccine. 2011;29:2649-52. 58. Piemonti L, Monti P, Allavena P, Leone BE, Caputo A, Di Carlo V. Glucocorticoids increase the endocytic activity of human dendritic cells. Int Immunol. 1999;11:1519-26. 59. Wollenberg A, Mommaas M, Oppel T, Schottdorf E-M, Günther S, Moderer M. Expression and Function of the Mannose Receptor CD206 on Epidermal Dendritic Cells in Inflammatory Skin Diseases. Journal of Investigative Dermatology. 2002;118(2):327-34. 60. Rodriguez A, Regnault A, Kleijmeer M, Ricciardi-Castagnoli P, Amigorena S. Selective transport of internalized antigens to the cytosol for MHC class I presentation in dendritic cells. Nature Cell Biology. 1999;1(6):362-8. 61. Knolle PA, Uhrig A, Hegenbarth S, Loser E, Schmitt E, Gerken G, et al. IL-10 down-regulates T cell activation by antigen-presenting liver sinusoidal endothelial cells through decreased antigen uptake via the mannose receptor and lowered surface expression of accessory molecules. Clin Exp Immunol. 1998;114:427-33. 62. Kang YS, Yamazaki S, Iyoda T, Pack M, Bruening SA, Kim JY, et al. SIGN-R1, a novel C-type lectin expressed by marginal zone macrophages in spleen, mediates uptake of the polysaccharide dextran. Int Immunol. 2003;15:177-86. 63. Lanoue A, Clatworthy MR, Smith P, Green S, Townsend MJ, Jolin HE, et al. SIGN-R1 contributes to protection against lethal pneumococcal infection in mice. J Exp Med. 2004;200:1383-93. 64. Koivusalo M, Welch C, Hayashi H, Scott CC, Kim M, Alexander T, et al. Amiloride inhibits macropinocytosis by lowering submembranous pH and preventing Rac1 and Cdc42 signaling. The Journal of Cell Biology. 2010;188(4):547-63. 65. Gu Z, Noss EH, Hsu VW, Brenner MB. Integrins traffic rapidly via circular dorsal ruffles and macropinocytosis during stimulated cell migration. The Journal of cell biology. 2011;193(1):61-70. 66. Schnatwinkel C, Christoforidis S, Lindsay MR, Uttenweiler-Joseph S, Wilm M, Parton RG, et al. The Rab5 Effector Rabankyrin-5 Regulates and Coordinates Different Endocytic Mechanisms. PLoS Biol. 2004;2(9). 67. Araki N, Hatae T, Yamada T, Hirohashi S. Actinin-4 is preferentially involved in circular ruffling and macropinocytosis in mouse macrophages: analysis by fluorescence ratio imaging. Journal of Cell Science. 2000;113(18):3329-40. 68. Garrett WS, Chen L-M, Kroschewski R, Ebersold M, Turley S, Trombetta S, et al. Developmental Control of Endocytosis in Dendritic Cells by Cdc42. Cell. 2000;102(3):325-34. 69. Racoosin EL, Swanson JA. Macropinosome maturation and fusion with tubular lysosomes in macrophages. The Journal of Cell Biology. 1993;121(5):1011-20. 70. Norbury CC, Hewlett LJ, Prescott AR, Shastri N, Watts C. Class I MHC presentation of exogenous soluble antigen via macropinocytosis in bone marrow macrophages. Immunity. 1995;3(6):783-91. 71. Dowrick P, Kenworthy P, McCann B, Warn R. Circular ruffle formation and closure lead to macropinocytosis in hepatocyte growth factor/scatter factor-treated cells. European journal of cell biology. 1993;61(1):44-53. 72. Rejman J, Oberle V, Zuhorn IS, Hoekstra D. Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochemical Journal. 2004;377(Pt 1):159-69. 73. Schroder U, Arfors KE, Tangen O. Stability of fluorescein labeled dextrans< i> in vivo and< i> in vitro. Microvascular Research. 1976;11:33-9. 74. Ohkuma S, Poole B. Fluorescence probe measurement of the intralysosomal pH in living cells and the perturbation of pH by various agents. Proceedings of the National Academy of Sciences. 1978;75(7):3327-31. 75. Sheedy FJ, Grebe A, Rayner KJ, Kalantari P, Ramkhelawon B, Carpenter SB, et al. CD36 coordinates NLRP3 inflammasome activation by facilitating intracellular nucleation of soluble ligands into particulate ligands in sterile inflammation. Nature immunology.14(8):812-20. 76. Gaudin Rl, Berre S, Cunha de Alencar B, Decalf Jrm, Schindler M, Gobert Fo-X, et al. Dynamics of HIV-Containing Compartments in Macrophages Reveal Sequestration of Virions and Transient Surface Connections. PLoS ONE.8(7). 77. Fiegl D, Kagebein D, Liebler-Tenorio EM, Weisser T, Sens M, Gutjahr M, et al. Amphisomal Route of MHC Class I Cross-Presentation in Bacteria-Infected Dendritic Cells. The Journal of Immunology. 2013;190(6):2791-806. 78. Zhang Z, Meng Y, Guo Y, He X, Li'u Q, Wan'g X, et al. Rehmannia glutinosa polysaccharide induces maturation of murine bone marrow derived Dendritic cells (BMDCs). International Journal of Biological Macromolecules. 2013;54:136-43. 79. Bauer R, Mezger M, Blockhaus C, Schmitt A-L, Kurzai O, Einsele H, et al. 40-O-[2-Hydroxyethyl] rapamycin modulates human dendritic cell function during exposure to Aspergillus fumigatus. Journal of Basic Microbiology. 2011;52(3):269-76. 80. Commisso C, Davidson SM, Soydaner-Azeloglu RG, Parker SJ, Kamphorst JJ, Hackett S, et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature.advance online publication. 81. Hope JC, Guzman E, Cubillos-Zapata C, Stephens SA, Gilbert SC, Prentice H, et al. Migratory sub-populations of afferent lymphatic dendritic cells differ in their interactions with Mycobacterium bovis Bacille Calmette Guerin. Vaccine. 2011;30(13):2357-67. 82. Gronwall A, Ingelman B. Dextran as a substitute for plasma. Nature. 1945;155:45-. 83. Vickery AL. The Fate of Dextran in Tissues of the Acutely Wounded: A Study of the Histologic Localization of Dextran in Tissues of Korean Battle Casualties*. The American Journal of Pathology. 1956;32. 84. Kulakov VN, Pimenova GN, Matveev VA, Sedov VV, Vasil'ev AE. Pharmacokinetic study of medicinal polymers: Models based on dextrans. Pharmaceutical Chemistry Journal. 1985;19:240-4. 85. Yamaoka T, Tabata Y, Ikada Y. Body distribution profile of polysaccharides after intravenous administration. Drug Delivery. 1993;1:75-82. 86. Mehvar R, Robinson MA, Reynolds JM. Molecular weight dependent tissue accumulation of dextrans: in vivo studies in rats. J Pharm Sci. 1994;83:1495-9. 87. Kaneo Y, Uemura T, Tanaka T, Kanoh S. Polysaccharides as drug carriers: biodisposition of fluorescein-labeled dextrans in mice. Biol Pharm Bull. 1997;20:181-7. 88. Howard JM, Teng CT, Loeffler RK, Johnsen A. Studies of dextrans of various molecular sizes. Annals of Surgery. 1956;143. 89. Rosenfeld EL, Lukomskaya IS. The splitting of dextran and isomaltose by animal tissues. Clinica chimica acta. 1957;2:105-14. 90. MacGregor E, Janecek S, Svensson B. Relationship of sequence and structure to specificity in the alfa-amylase family of enzymes. Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular Enzymology. 2001;1546:1-20. 91. Artimo P, Jonnalagedda M, Arnold K, Baratin D, Csardi G, de Castro E, et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Research. 2012;40(W1):W597-W603. 92. Schomburg I, Chang A, Placzek S, Söhngen C, Rother M, Lang M, et al. BRENDA in 2013: integrated reactions, kinetic data, enzyme function data, improved disease classification: new options and contents in BRENDA. Nucleic Acids Research. 2013;41(Database issue):D764-72. 93. Cargill WH, Bruner HD. Metabolism of C-14 labeled dextran in the mouse. J Pharmacol. 1951;103. 94. Gray I. Metabolism of plasma expanders studied with carbon-14-labeled dextran. American Journal of Physiology--Legacy Content. 1953;174. 95. Schlesinger LS. Macrophage phagocytosis of virulent but not attenuated strains of Mycobacterium tuberculosis is mediated by mannose receptors in addition to complement receptors. J Immunol. 1993;150:2920-30. 96. Kang PB, Azad AK, Torrelles JB, Kaufman TM, Beharka A, Tibesar E, et al. The human macrophage mannose receptor directs Mycobacterium tuberculosis lipoarabinomannan-mediated phagosome biogenesis. J Exp Med. 2005;202:987-99. 97. Astarie-Dequeker C, N'Diaye EN, Le Cabec V, Rittig MG, Prandi J, Maridonneau-Parini I. The mannose receptor mediates uptake of pathogenic and nonpathogenic mycobacteria and bypasses bactericidal responses in human macrophages. Infection and immunity. 1999;67. 98. Nguyen DG, Hildreth JEK. Involvement of macrophage mannose receptor in the binding and transmission of HIV by macrophages. European journal of immunology. 2003;33:483-93. 99. Milone MC, Fitzgerald-Bocarsly P. The Mannose Receptor Mediates Induction of IFN-О± in Peripheral Blood Dendritic Cells by Enveloped RNA and DNA Viruses. The Journal of Immunology. 1998;161(5):2391-9. 100. Bandivdekar AH. P1.031 CD4 Independent Binding of HIV to Human Mannose Receptor on Vaginal Epithelial Cells and Sperm. Sexually Transmitted Infections.89(Suppl 1):A83-A. 101. Crespo H, Reina R, Glaria I, Ramírez H, de Andrés X, Jáuregui P, et al. Identification of the ovine mannose receptor and its possible role in Visna/Maedi virus infection. Vet Res. 2011;42. 102. Crespo H, Jauregui P, Glaria I, Sanjosé L, Polledo L, García-Marín JF, et al. Mannose receptor may be involved in small ruminant lentivirus pathogenesis. Veterinary research. 2012;43(1):1-6. 103. Marodi L, Schreiber S, Anderson DC, MacDermott RP, Korchak HM, Johnston RB. Enhancement of macrophage candidacidal activity by interferon-gamma. Increased phagocytosis, killing, and calcium signal mediated by a decreased number of mannose receptors. Journal of Clinical Investigation. 1993;91(6):2596-601. 104. Newman SL, Holly A. Candida albicans Is Phagocytosed, Killed, and Processed for Antigen Presentation by Human Dendritic Cells. Infection and immunity. 2001;69(11):6813-22. 105. Reading PC, Miller JL, Anders EM. Involvement of the Mannose Receptor in Infection of Macrophages by Influenza Virus. Journal of virology. 2000;74(11):5190-7. 106. Miller JL, deWet BJM, Martinez-Pomares L, Radcliffe CM, Dwek RA, Rudd PM, et al. The Mannose Receptor Mediates Dengue Virus Infection of Macrophages. PLoS Pathog. 2008;4(2). 107. Op den Brouw ML, Binda RS, Geijtenbeek TBH, Janssen HLA, Woltman AM. The mannose receptor acts as hepatitis B virus surface antigen receptor mediating interaction with intrahepatic dendritic cells. Virology. 2009;393(1):84-90. 108. Everts B, Hussaarts L, Driessen NN, Meevissen MHJ, Schramm G, Ham AJvd, et al. Schistosome-derived omega-1 drives Th2 polarization by suppressing protein synthesis following internalization by the mannose receptor. The Journal of experimental medicine. 2012;209(10):1753-67. 109. Hofmann H, Pohlmann S. DC-SIGN: Access Portal for Sweet Viral Killers. Cell Host & Microbe. 2011;10(1):5-7. 110. de Witte L, de Vries RD, van der Vlist M, YГјksel S, Litjens M, de Swart RL, et al. DC-SIGN and CD150 have distinct roles in transmission of measles virus from dendritic cells to T-lymphocytes. PLoS pathogens. 2008;4(4). 111. Schulert GS, Allen L-AH. Differential infection of mononuclear phagocytes by Francisella tularensis: role of the macrophage mannose receptor. Journal of Leukocyte Biology. 2006;80(3):563-71. 112. Zhang P, Skurnik M, Zhang S-S, Schwartz O, Kalyanasundaram R, Bulgheresi S, et al. Human dendritic cell-specific intercellular adhesion molecule-grabbing nonintegrin (CD209) is a receptor for Yersinia pestis that promotes phagocytosis by dendritic cells. Infection and immunity. 2008;76(5):2070-9. 113. Wilson ME, Pearson RD. Roles of CR3 and mannose receptors in the attachment and ingestion of Leishmania donovani by human mononuclear phagocytes. Infection and immunity. 1988;56(2):363-9. 114. Ueno N, Wilson ME. Receptor-mediated phagocytosis of< i> Leishmania</i>: implications for intracellular survival. Trends in parasitology. 2012;28(8):335-44. 115. Tailleux L, Schwartz O, Herrmann JL, Pivert E, Jackson M, Amara A, et al. DC-SIGN is the major Mycobacterium tuberculosis receptor on human dendritic cells. J Exp Med. 2003;197:121-7. 116. Sarkar R, Mitra D, Chakrabarti S. HIV-1 Gp120 Protein Downregulates Nef Induced IL-6 Release in Immature Dentritic Cells through Interplay of DC-SIGN. PLoS ONE. 2013;8(3). 117. Cambi A, Gijzen K, de Vries IJM, Torensma R, Joosten B, Adema GJ, et al. The C-type lectin DC-SIGN (CD209) is an antigen-uptake receptor for Candida albicans on dendritic cells. European journal of immunology. 2003;33(2):532-8. 118. Londrigan SL, Tate MD, Brooks AG, Reading PC. Cell-surface receptors on macrophages and dendritic cells for attachment and entry of influenza virus. Journal of Leukocyte Biology. 2012;92:97-106. 119. Hillaire MLB, Nieuwkoop NJ, Boon ACM, de Mutsert G, Vogelzang-van Trierum SE, Fouchier RAM, et al. Binding of DC-SIGN to the Hemagglutinin of Influenza A Viruses Supports Virus Replication in DC-SIGN Expressing Cells. PLoS ONE.8(2). 120. Marzi A, Gramberg T, Simmons G, Möller P, Rennekamp AJ, Krumbiegel M, et al. DC-SIGN and DC-SIGNR Interact with the Glycoprotein of Marburg Virus and the S Protein of Severe Acute Respiratory Syndrome Coronavirus. Journal of virology. 2004;78(21):12090-5. 121. Zhang Y, Buckles E, Whittaker GR. Expression of the C-type lectins DC-SIGN or L-SIGN alters host cell susceptibility for the avian coronavirus, infectious bronchitis virus. Veterinary Microbiology. 2012;157(3–4):285-93. 122. Goncalves A-R, Moraz M-L, Pasquato A, Helenius A, Lozach P-Y, Kunz S. Role of DC-SIGN in Lassa Virus Entry into Human Dendritic Cells. Journal of virology.87(21):11504-15. 123. Martinez MG, Bialecki MA, Belouzard S, Cordo SM, Candurra NlA, Whittaker GR. Utilization of human DC-SIGN and L-SIGN for entry and infection of host cells by the New World arenavirus, Junín virus. Biochemical and Biophysical Research Communications. 2013;441(3):612-7. 124. Lozach P-Y, Lortat-Jacob H, Lavalette ADLD, Staropoli I, Foung S, Amara A, et al. DC-SIGN and L-SIGN Are High Affinity Binding Receptors for Hepatitis C Virus Glycoprotein E2. Journal of Biological Chemistry. 2003;278(22):20358-66. 125. Tassaneetrithep B, Burgess TH, Granelli-Piperno A, Trumpfheller C, Finke J, Sun W, et al. DC-SIGN (CD209) Mediates Dengue Virus Infection of Human Dendritic Cells. The Journal of experimental medicine. 2003;197(7):823-9. 126. Hottz ED, Oliveira MF, Nunes PCG, Nogueira RMR, Valls-de-Souza R, Da Poian AT, et al. Dengue induces platelet activation, mitochondrial dysfunction and cell death through mechanisms that involve DC-SIGN and caspases. Journal of Thrombosis and Haemostasis.11(5):951-62. 127. Davis CW, Nguyen H-Y, Hanna SL, Sánchez MD, Doms RW, Pierson TC. West Nile Virus Discriminates between DC-SIGN and DC-SIGNR for Cellular Attachment and Infection. Journal of virology. 2006;80(3):1290-301. 128. Barkhash AV, Perelygin AA, Babenko VN, Brinton MA, Voevoda MI. Single nucleotide polymorphism in the promoter region of the CD209 gene is associated with human predisposition to severe forms of tick-borne encephalitis. Antiviral research. 2012;93(1):64-8. 129. Johnson TR, McLellan JS, Graham BS. Respiratory Syncytial Virus Glycoprotein G Interacts with DC-SIGN and L-SIGN To Activate ERK1 and ERK2. Journal of virology. 2012;86(3):1339-47. 130. Halary F, Amara A, Lortat-Jacob H, Messerle M, Delaunay T, Houlès C, et al. Human Cytomegalovirus Binding to DC-SIGN Is Required for Dendritic Cell Infection and Target Cell trans-Infection. Immunity. 2002;17(5):653-64. 131. Rappocciolo G, Jenkins FJ, Hensler HR, Piazza P, Jais M, Borowski L, et al. DC-SIGN is a receptor for human herpesvirus 8 on dendritic cells and macrophages. The Journal of Immunology. 2006;176(3):1741-9. 132. Rappocciolo G, Hensler HR, Jais M, Reinhart TA, Pegu A, Jenkins FJ, et al. Human herpesvirus 8 infects and replicates in primary cultures of activated B lymphocytes through DC-SIGN. Journal of virology. 2008;82(10):4793-806. 133. Alvarez CP, Lasala F, Carrillo J, Muniz O, Corbi AL, Delgado R. C-type lectins DC-SIGN and L-SIGN mediate cellular entry by Ebola virus in cis and in trans. J Virol. 2002;76:6841-4. 134. Appelmelk BJ, van Die I, van Vliet SJ, Vandenbroucke-Grauls CM, Geijtenbee'k TB, van Kooyk Y. Cutting edge: carbohydrate profiling identifies new pathogens that interact with dendritic cell-specific ICAM-3-grabbing nonintegrin on dendritic cells. J Immunol. 2003;170:1635-9. 135. Caparros E, Serrano D, Puig-Kroger A, Riol L, Lasala F, Martinez I, et al. Role of the C-type lectins DC-SIGN and L-SIGN in Leishmania interaction with host phagocytes. Immunobiology. 2005;210(2-4):185-93. 136. Lefevre L, Lugo-Villarino G, Meunier E, Valentin A, Olagnier D, Authier Hl, et al. The C-type Lectin Receptors Dectin-1, MR, and SIGNR3 Contribute Both Positively and Negatively to the Macrophage Response to Leishmania infantum. Immunity. 2013;38(5):1038-49. 137. van Die I, van Vliet SJ, Nyame AK, Cummings RD, Bank CMC, Appelmelk B, et al. The dendritic cell-specific C-type lectin DC-SIGN is a receptor for Schistosoma mansoni egg antigens and recognizes the glycan antigen Lewis x. Glycobiology. 2003;13(6):471-8. 138. Klimstra WB, Nangle EM, Smith MS, Yurochko AD, Ryman KD. DC-SIGN and L-SIGN Can Act as Attachment Receptors for Alphaviruses and Distinguish between Mosquito Cell- and Mammalian Cell-Derived Viruses. Journal of virology. 2003;77(22):12022-32. 139. Iyori M, Ohtani M, Hasebe A, Totsuka Y, Shibata K-i. A role of the Ca2+ binding site of DC-SIGN in the phagocytosis of E. coli. Biochemical and Biophysical Research Communications. 2008;377(2):367-72. 140. Bloem K, Garcia-Vallejo JJ, Vuist IM, Cobb BA, van Vliet SJ, van Kooyk Y. Interaction of the Capsular Polysaccharide A from Bacteroides fragilis with DC-SIGN on Human Dendritic Cells is Necessary for Its Processing and Presentation to T Cells. Frontiers in Immunology. 2013;4. 141. Koppel EA, Ludwig IS, Hernandez MS, Lowary TL, Gadikota RR, Tuzikov AB, et al. Identification of the mycobacterial carbohydrate structure that binds the C-type lectins DC-SIGN, L-SIGN and SIGNR1. Immunobiology. 2004;209:117-27. 142. Taylor PR, Brown GD, Herre J, Williams DL, Willment JA, Gordon S. The Role of SIGNR1 and the ОІ-Glucan Receptor (Dectin-1) in the Nonopsonic Recognition of Yeast by Specific Macrophages. The Journal of Immunology. 2004;172(2):1157-62. 143. Koppel EA, Wieland CW, van den Berg VCM, Litjens M, Florquin S, van Kooyk Y, et al. Specific ICAM-3 grabbing nonintegrin-related 1 (SIGNR1) expressed by marginal zone macrophages is essential for defense against pulmonary Streptococcuspneumoniae infection. European journal of immunology. 2005;35(10):2962-9. 144. Koppel EA, Litjens M, van den Berg VC, van Kooyk Y, Geijtenbeek TBH. Interaction of SIGNR1 expressed by marginal zone macrophages with marginal zone B cells is essential to early IgM responses against Streptococcus pneumoniae. Molecular immunology. 2008;45(10):2881-7. 145. Tanne A, Ma B, Boudou F, Tailleux L, Botella H, Badell E, et al. A murine DC-SIGN homologue contributes to early host defense against Mycobacterium tuberculosis. J Exp Med. 2009;206:2205-20. 146. Pohlmann S, Soilleux EJ, Baribaud F, Leslie GJ, Morris LS, Trowsdale J, et al. DC-SIGNR, a DC-SIGN homologue expressed in endothelial cells, binds to human and simian immunodeficiency viruses and activates infection in trans. Proc Natl Acad Sci U S A. 2001;98:2670-5. 147. Gardner JP, Durso RJ, Arrigale RR, Donovan GP, Maddon PJ, Dragic T, et al. L-SIGN (CD 209L) is a liver-specific capture receptor for hepatitis C virus. Proceedings of the National Academy of Sciences. 2003;100(8):4498-503. 148. Van Liempt E, Imberty A, Bank CM, Van Vliet SJ, Van Kooyk Y, Geijtenbeek TB, et al. Molecular basis of the differences in binding properties of the highly related C-type lectins DC-SIGN and L-SIGN to Lewis X trisaccharide and Schistosoma mansoni egg antigens. J Biol Chem. 2004;279:33161-7. 149. Simmons G, Reeves JD, Grogan CC, Vandenberghe LH, Baribaud Fdr, Whitbeck JC, et al. DC-SIGN and DC-SIGNR Bind Ebola Glycoproteins and Enhance Infection of Macrophages and Endothelial Cells. Virology. 2003;305(1):115-23. 150. Hunger RE, Sieling PA, Ochoa MT, Sugaya M, Burdick AE, Rea TH, et al. Langerhans cells utilize CD1a and langerin to efficiently present nonpeptide antigens to T cells. Journal of Clinical Investigation. 2004;113(5):701-8. 151. de Witte L, Nabatov A, Pion M, Fluitsma D, de Jong MAWP, de Gruijl T, et al. Langerin is a natural barrier to HIV-1 transmission by Langerhans cells. Nature Medicine. 2007;13(3):367-71. 152. de Jong MAWP, Vriend LEM, Theelen B, Taylor ME, Fluitsma D, Boekhout T, et al. C-type lectin Langerin is a ОІ-glucan receptor on human Langerhans cells that recognizes opportunistic and pathogenic fungi. Molecular immunology. 2010;47(6):1216-25. 153. van der Vlist M, Geijtenbeek TBH. Langerin functions as an antiviral receptor on Langerhans cells. Immunology and Cell Biology. 2011;88(4):410-5. 154. Liu B, Wan'g M, Wan'g X, Zhao D, Liu D, Liu J, et al. Liver Sinusoidal Endothelial Cell Lectin Inhibits CTL-Dependent Virus Clearance in Mouse Models of Viral Hepatitis. The Journal of Immunology.190(8):4185-95. 155. Gramberg T, Hofmann H, Möller P, Lalor PF, Marzi A, Geier M, et al. LSECtin interacts with filovirus glycoproteins and the spike protein of SARS coronavirus. Virology. 2005;340(2):224-36. 156. Powlesland AS, Fisch T, Taylor ME, Smith DA, Tissot B, Dell A, et al. A Novel Mechanism for LSECtin Binding to Ebola Virus Surface Glycoprotein through Truncated Glycans. Journal of Biological Chemistry. 2008;283(1):593-602. 157. L'i Y, Hao B, Kuai X, Xing G, Yang J, Chen J, et al. C-type lectin LSECtin interacts with DC-SIGNR and is involved in hepatitis C virus binding. Molecular and Cellular Biochemistry. 2009;327(1-2):183-90. 158. Shimojima M, Ströher U, Ebihara H, Feldmann H, Kawaoka Y. Identification of Cell Surface Molecules Involved in Dystroglycan-Independent Lassa Virus Cell Entry. Journal of virology. 2012;86(4):2067-78. 159. Shkurupy VA. Tuberculosis-induced granulomatosis. Cytophysiology and targeted therapy [in Russian]. 2007. 160. Elkington PT, D'Armiento JM, Friedland JS. Tuberculosis immunopathology: the neglected role of extracellular matrix destruction. Science translational medicine. 2011;3(71):71ps6-ps6. 161. Dartois V. The path of anti-tuberculosis drugs: from blood to lesions to mycobacterial cells. Nature Reviews Microbiology. 2014;12(3):159-67. 162. Shkurupy VA, Kurunov JN, Arkhipov SA. [Antibacterial efficiency of prolonged isoniazid formulation in experiment] [In Russian]. Problemy tuberkuleza. 1997(2):54-6. 163. Mantegazza AR, Savina A, Vermeulen M, Pérez L, Geffner J, Hermine O, et al. NADPH oxidase controls phagosomal pH and antigen cross-presentation in human dendritic cells. Blood. 2008;112(12):4712-22. 164. Shkurupy VA, Selyatitskaya VG, Tsyrendorzhiev DD, Pal'chikova NA, Kurilin VV, Travin MA, et al. Effects of modified amphotericin in experimental systemic candidiasis. Bull Exp Biol Med. 2007;143:392-4. 165. Polyakova IN, Ginak AI, Moskvichev BV, Shitikova GS. Chemical modification of polysacharides with rimantadine and the properties of derived product [in Russian]. Isvestia VolgGTU. 2011;2 (75)(8):99-102. 166. Potapova OV, Shkurupiy VA. Effect of dialdehyde dextran on structural changes in the liver and lungs during chronic BCG granulomatosis. Bull Exp Biol Med. 2008;146:861-3. 167. Potapova OV, Shkurupiy VA, Sharkova TV, Troitskiy AV, Lusgina NG, Shestopalov AM. Preventive Efficacy of Oxidized Dextran and Pathomorphological Processes in Mouse Lungs in Avian Influenza A/H5N1. Bulletin of Experimental Biology and Medicine. 2011;150:707-10. 168. Shkurupy V, Guseva E, Potapov'a O, Nadeev A. Morphological Changes in the Brain of Mice with Systemic Candidiasis Treated with Composition of Amphotericin B and Oxidized Dextran. Bulletin of Experimental Biology and Medicine. 2011:1-4. 169. Shkurupy VA, Kozyaev MA, Potapov'a OV. Morphological Study of the Efficiency of Isoniazid and Dialdehyde Dextran Composition in the Treatment of Mice with BCG Granulomatosis. Bulletin of experimental biology and medicine. 2008;146:853-6. 170. Ryan AA, Wozniak TM, Shklovskaya E, O' Donnell MA, Groth BFdS, Britton WJ, et al. Improved Protection against Disseminated Tuberculosis by Mycobacterium bovis Bacillus Calmette-Guerin Secreting Murine GM-CSF Is Associated with Expansion and Activation of APCs. The Journal of Immunology. 2007;179(12):8418-24. 171. Tkachev VO, Kolesnikova OP, Troitskii AV, Shkurupy VA. Effect of oxidized dextrans on NO synthase and arginase activities of mouse macrophages. Bull Exp Biol Med. 2008;146:83-5. 172. Tkachev VO, Zaikovskaya MV, Troitsky AV, Luzgina NG, Shkurupy VA. The effect of oxidized dextrans on generation of reactive oxygen species by murine peritoneal exudate phagocytes. Biochemistry (Moscow) Supplement Series B: Biomedical Chemistry. 2012;6(2):144-8. 173. Pustylnikov S. Dextrans as Ligands of Mannose Receptor and DC-SIGN Family Receptors - A Novel Means of Modulating Th1/Th2 Immune Responses (Abstract Book). Annals of Allergy, Asthma & Immunology. 2012;109:A108,-P248. 174. Pustylnikov S, DuBuske L. Inhibiting the binding of allergens having glycosylated structures by dextrans as a potential immunomodulatory strategy. European Academy of Allergy and Clinical Immunology and World Allergy Organization Congress, 2013, Milan, Italy, June 22-26, 2013; 2013; 2013. p. 885. 175. Royer P-J, Emara M, Yang C, Al-Ghouleh A, Tighe P, Jones N, et al. The Mannose Receptor Mediates the Uptake of Diverse Native Allergens by Dendritic Cells and Determines Allergen-Induced T Cell Polarization through Modulation of IDO Activity. The Journal of Immunology. 2010;185(3):1522-31. 176. Emara M, Royer P-J, Mahdavi J, Shakib F, Ghaemmaghami AM. Retagging Identifies Dendritic Cell-specific Intercellular Adhesion Molecule-3 (ICAM3)-grabbing Non-integrin (DC-SIGN) Protein as a Novel Receptor for a Major Allergen from House Dust Mite. Journal of Biological Chemistry. 2011;287(8):5756-63. 177. Salazar F, Sewell HF, Shakib F, Ghaemmaghami AM. The role of lectins in allergic sensitization and allergic disease. Journal of Allergy and Clinical Immunology. 2013;132(1):27-36. 178. Pustylnikov S, Zavjalov E, Litvinova E. Dextran affects the immune response of BALB/c mice to intranasal challenge with inactivated mycobacterium H37Rv - Abstract in Russian. Russian Laboratory Animal Science Association Conference, 2013, Novosibirsk, Russia, September 25-28; 2013; 2013. p. 39. 179. Pustylnikov S.V MAG, Durymanov A.G. Dextran Extends an Average Length of Life of Mice after Avian Influenza H5N1 Infection. General Meeting of the American Society of Microbiology 2014; 2014 May 17-20, 2014; Boston; 2014. 180. Royce RA, Sena A, Cates Jr W, Cohen MS. Sexual transmission of HIV. New England Journal of Medicine. 1997;336(15):1072-8. 181. Wu L. Biology of HIV mucosal transmission. Current Opinion in HIV and AIDS. 2008;3(5):534. 182. Arrighi J-Fo, Pion M, Garcia E, van Kooyk Y, Geijtenbeek TB, Piguet V. DC-SIGN-mediated Infectious Synapse Formation Enhances X4 HIV-1 Transmission from Dendritic Cells to T Cells. The Journal of experimental medicine. 2004;200(10):1279-88. 183. Hodges A, Sharrocks K, Edelmann M, Baban D, Moris A, Schwartz O, et al. Activation of the lectin DC-SIGN induces an immature dendritic cell phenotype triggering Rho-GTPase activity required for HIV-1 replication. Nature Immunology. 2007;8(6):569-77. 184. Avota E, Gulbins E, Schneider-Schaulies S. DC-SIGN mediated sphingomyelinase-activation and ceramide generation is essential for enhancement of viral uptake in dendritic cells. PLoS pathogens. 2011;7(2):e1001290. 185. Gringhuis SI, van der Vlist M, van den Berg LM, den Dunnen J, Litjens M, Geijtenbeek TBH. HIV-1 exploits innate signaling by TLR8 and DC-SIGN for productive infection of dendritic cells. Nature Immunology. 2010;11(5):419-26. 186. Este JA, Telenti A. HIV entry inhibitors. The Lancet. 2007;370(9581):81-8. 187. Feinberg H, Mitchell DA, Drickamer K, Weis WI. Structural basis for selective recognition of oligosaccharides by DC-SIGN and DC-SIGNR. Science. 2001;294(5549):2163-6. 188. Wen J, Pang L, Estes D. The roles of combination of dextran with gp120 on human macrophage immune response in HIV infection. The Journal of Immunology. 2012;188:168.16. 189. Chehimi J, Luo Q, Azzoni L, Shawver L, Ngoubilly N, June R, et al. HIV-1 transmission and cytokine-induced expression of DC-SIGN in human monocyte-derived macrophages. Journal of Leukocyte Biology. 2003;74(5):757-63. 190. Frohlich JA, Grobler AC, Baxter C, Mansoor LE, Kharsany AB, Sibeko S, et al. Effectiveness and safety of tenofovir gel, an antiretroviral microbicide, for the prevention of HIV infection in women. Science (New York, NY). 2010;329(5996):1168-74. 191. Neff CP, Kurisu T, Ndolo T, Fox K, Akkina R. A topical microbicide gel formulation of CCR5 antagonist maraviroc prevents HIV-1 vaginal transmission in humanized RAG-hu mice. PLoS ONE. 2011;6(6):e20209. 192. Ingelman B. Dextran and its use as a plasma substitute. Acta
|