
Биологиялық жүйелерден электр тогы өткен кездегі тағайындалған заңдылықтары
Тірі клетка мен ткандерден электірлік тогын өткізген кезде, ток күші тұрақты болып қалмайды, потенциал берілгеннен кейін токтың шамасы бірден кеміп сонында бастапқы мәнінің деңгейінен де төмен болады. Себебі тұрақты ток биологиялық системадан өткен кезде онда керібағытта белгілі бір шекке дейін өсетін э.қ.к. пайда болады. Осы қарама – қарсы пайда болған э.қ.к.-ті табу (білу) үшін кернеу көзін электродтардан тез ажыратып бірден гальвонометрге жалғаса кері бағытты ток жүріп, ол уақыт бойынша азаяды. Қарама – қарсы ә.к.к. пайда болуы Ом заңынан ауытқуды көрсетеді. Металдарда дәлме – дәл орындалатын Ом заңы, тірі организмге сол күйінде орындалмайды. Өткізгішке берілетін кернеу тұрақты болса онда өтетін ток күші де тұрақты болады бірақ бұл заң тірі организде орындалмайды Осы жағдай үшін Ом заңы былай жазылады. Тірі биологиялық объект үшін Ом заңы былайша өрнектеледі Мұндағы P = g (t) – уақыт функциясы- поляризация қозғаушы күші. Байқалған құбылыс электролит ертіндісінде болатын құбылысқа ұқсас, ал оған поляризация құбылысы сәйкес келеді. Яғни тұрақты ток өткенде кері таңбалы иондардың шоғырлану нәтижесінде қосымша зарядтар пайда болады. Биологиялық системада – ток күшінің өзгеруі олардың ток күшін поляризациялай алатын қасиеттерінің бар екендігін көрсетеді. Тірі организмен ток өтіп жатыр делік, оған берілген кернеу өзгермесе де, одан өтетін ток өзгереді, яғни ток күші кемиді. Ток күшінің кемуі тканьде болып жатқан поляризацияға байланысты. Атап айтқанда, тканьдардың сиымдылық, диэлектрлік қасиеттерінен ток күші азаяды. Байланысқан зарядтардың электр өрісі әсерінен қозғалуын сөйтіп электр қозғаушы күшінің пайда болуын поляризация деп атайды. Диэлектриктер поляризациясы электрондық – иондық - дипольдік - макроструктуралық – беттік және электролиттік болып бөлінеді 1. Электрондық – поляризация деп өз орбитасында қозғалып жүрген электроның он зарядталған ядроға қарағанда ығысуын айтады. Бұл ығысудын нәтижесінде атом бағыты бар дипольге ұқсап кетеді. Электрондық поляризацияның пайда болу уақытын релаксация уақыты деп атайды Ол τ = 10-16 ÷ 10-14 с 2. Иондық поляризация деп ионның кристалл торына қарағанда ығысуын айтады. Бұл кездегі релаксация уақыты τ = 10-14 ÷ 10-12 с тен болады. 3. Дипольдық поляризация деп полярлық молекулалардың электр өрісінде орналасуын айтады. Бұл поляризацияның релаксация уақыт τ = 10-13 ÷ 10-7 с тең. 4. Макроструктуралық поляризация заттардың электр қасиеттерінің әр түрлілігінен пайда болады. Электр өткізгіштігі әр түрлі бірнеше қабаттан тұратын заттын бетінде осы поляризация пайда болды. Бұл поляразацияда релаксация уақыты τ = 10-8 ÷ 10-3 с. 5. Беттік поляризация - екі электр қабаттан тұратын беттерде пайда болады τ=10-3 ÷ 100 с. 6. Электролиттік поляризация – электролиттен ток жүріп жатқан оған батырылған екі электродтың арасында пайда болады. Бұл поляризацияның реакциясы уақыты τ = 10-4÷ 10-2 с. Осы айтылған поляризация ткандер мен клеткаларда, жалпы биологиялық объектілерде байқалады. Бұл поляризациялар заттардың диэлектрлік өнімділігімен сипатталады. (Кесте-2) Тірі системада жинақталған электр мөлшері статикалық сиымдылықтың шамаларымен ғана анықталмай оған өте үлкен шамадағы салыстырмалы поляризацияланған сиымдылық қосылады.
Кесте-2 Биологиялық жүйелердің диэлектрлік өтімділіктерінің тұрақты жиіліктегі өзгерісі
Статикалық сиымдылық жазық конденсатордың теңдеуімен есептеледі.
Поляризцияланған сиымдылықтың шамасы ток күшінің бастапқы және соңғы мәндеріне байланысты болып мына теңдеу бойынша есептеледі.
Ср – поляризациялық сиымдылық R – кедергі, J- ток күші J0 – ток күшінің бастапқы мәні J – ток күшінің соңғы мәні
Әртүрлі биологиялық обектілердің тұрақты ток өткен кездегі өлшенген поляризацияланған сиымдылығы өте үлкен шамаларға ие болады, 0,1мкф /см2 тан 10 ÷ 40мкф / см2 - ке жетеді. Жоғары поляризацияланған сиымдылық – тірі бұзылмаған клеткалардың сипаттамалық қасиеті болып саналады. Тірі клетка мен ткандердің қай жерінде поляризация құбылысы пайда болады. Поляризация – электролиттің ертіндісінде оған түсірілген (ендірілген) электродтарда пайда болады. Олардың иондық адсорбция құбылысы қарама – қарсы бағытта пайда болатын ә.к.к. туғызады. Электр потенциалдары қойылған кезде пайда болатын поляризация құбылысын басқа да модельдерде көруге болады. Мысалы кристалдарда, диэлектриттерде. Бұл жерде поляризацияның 2 түрін ажыратуға болады. 1. Беттік. 2. Көлемдік. 1. Беттік поляризация (кальцит кристалында) - берілген жұқа беттік қабатта иондардың электродтың жанында жиналуынан пайда болады. Егер кристалдың беттік қабатын тегістесек поляризация жойылады. 2. Көлемдік поляризация (мыс -кварц кристалында) берілген кристалдың барлығы қалындық бойы зарядтардың шоғырлануынан пайда болады. Бұл кезде беттік жұқа қабатты алып тастасақта поляризация жоғалмайды. Тірі клеткалар мен ткандерде поляризация құбылысын бос еркін иондар туғызады, ал поляризациялық сиымдылық клетканың бетінде жиналып шоғырланады. Биологиялық жүйе арқылы ток өтуі кезінде пайда болатын кейбір заңдылықтарды қарастырайық. Тұрақты токта биологиялық объектілердің кедергісін анықтау поляризация құбылысының нәтижесінде өте қиын. Сонымен қатар тірі клетка арқылы электр тогы өткенде протпоплазманың дезинтерграция құбылысы байқалады, ал бұл клетканың бұлінуіне және оның электрөткізгіштігінің тез өсуіне әкеп соғады. Электролит ертіндісіндегі кедергіні анықтау үшін поляризация құбылысынан құтылу үшін Кольрауш айнымалы токты пайдалануды ұсынды. Айнымалы токпен тірі ткандерде жұмыс істеу барысында, арнаулы көпірі бар компенсациялық схемалар пайдалынылады, соның нәтижесінде мына заңдылықтар тағайындалған. 1. Aйнымалы токтағы биологиялық объектінің кедергісі, тұрақты токқа қарағанда кем болады R<R 2. Егер токтың шамасы физиологиялық нормадан аспаса онда кедергі токтың шамасына байланысты болмайды. 3. Арнаулы көпір схемасының айнымалы иініне, Омдық кедергі мен қатар Сиымдылықты пайдалану өте тиімді балады. 4. Биологиялық объектілердің физиологиялық күйлері өзгермесе, берілген жилікте олардың кедергілері тұрақты болады. 5. Биологиялық системаның физиологиялық күйі өзгергенде, оның кедергісі де өзгереді, ал жансыздана бастағанда кедергі кемиді. 6. Тірі клетка мен ткандердің электрөткізгіштігін өлшеу, бір тәжірбиенің өзін әр түрлі жиліктерде қайталауға мүмкіндіктің болуы оның маңыздылығын арттыра түсті. Электролит ертіндісі арқылы айнымалы токты өткізгенде олардың электрөткізгіштігі жилікке тәуелді болмай бірдей мәндерге ие болады (1-10Гц деиін). Биологиялық объектімен, мысалы қанмен жұмыс істегенде, Гебер мынандай қызғылықты заңдылықты байқады, яғни жоғарғы жиліктегі (107 Гц) электрөткізгіштік – төменгі жиліктегі электрөткізгіштікке қарағанда өте жоғары болады. Бұл задылық кейінгі тәжірибелерменде дәлелденді. Яғни биологиялық объектілердің электрөткізгіштігі жилік өскен сайын белгілі бір максимум шамаға дейін өседі. 4.2.-суретте – Филлипсон тәжірибесіне сәйкес (1920) бұлшық ет кедергісінің айнымалы ток жилігіне тәуелділік графигі берілген.
4.2.-сурет.Бұлшықет кедергісінің тоқ жиілігінің өсуіне байланыстылығы.
Кейінгі зерттеулерде көрсеткендей мұндай тәуелділік олардың кедергілерінің абсолют мәндеріне байланысты емес кез келген тірі клетка мен ткандерге тән болып шықты. Электрөткізгіштіктің дисперсиялық зонасы кәдімгі 102 –108Гц аралық интервалында өзгереді. Көптеген биологиялық объект үшін электрөткізгіштіктің максималдық мәні 106Гц жилікте байқалады. Төменгі жиліктегі тұрақты ток кезінде тірі биологиялық жүйедегі электрөткізгіштіктің дисперсиясының пайда болуы, электрөткізгіштік поляризациясымен байланысты – жилік өскен сайын поляризациялық құбылыстар аз байқалады. Электркізгіштік дисперсиясы – поляризация құбылысына қаблеттігі сияқты тек тірі клеткаларға ғана тән. Тәжірибе нәтижелері көрсеткендей – Дисперсия қисығының крутизна биіктігі - ткань жансыздана бастаған сайын кемиді, яғни төменгі кедергі өзгермей тұрақты болып қалады. Остергаут – ткань жансыздана бастаған кезде оның төменгі жиліктегі кедергісінің кеми беретіндігін көрсеткен. Одан кейінгі зерттеулерде төменгі жилікте кедергілерді өлшеу - биологиялық объектілердің күйлерін сипаттау үшін қолданыла бастады. Бұл критерий - трансплантацияланатын ткандердің қаншалықты жанды екендігін анықтау мен бағалауда пайдалануда. Бірақ кедергінің абсалют мәнін алу өте қиын, себебі ол шама электродтардың ауданына, олардың ара қашықтығына, ткандердің элетродтардың жанында тығыз орналасқанына және объектінің қалындығына т.б. байланысты болады. 4.3.-суретте - өсімдік ұлпасының жансызданып бара жатқан кезіндегі өзгерісінің электрөткізгіштік дисперциясы берілген.
4.3.-сурет. Өсімдік ұлпасының кедергісінің (R) тоқ жиілігіне байланыстылығы. 1. Қалыпты жағдай; 2.50˚С-та 2 мин қыздыру; 3. 50˚С-та 4мин қыздыру; 4. 20 мин қайнату кезінде.
Биологиялық объектінің физиологиялық күйлерін бағалаудың ең бір сенімді жолы - электрөткізгіштік дисперсиясының (крутизна) тік биіктігін сипаттау. Дисперсияның тік толық қисығын алу – техникалық жағынан қиын Б.Н. Тарусов - дисперсияның биіктік қисықтығын сипаттау үшін – төменгі жиліктегі кедергі шамасының – жоғарғы жиліктегі кедергі шамасына қатына сын алуды ұсынды. Кедергіні өлшеу үшін 106 Гц және 104Гц жиліктер таңдалып алынды, себебі көптеген жағайда 106 Гц жилікте биологиялық объектінің максимал кедергісі байқалады, ал 104 Гц жилікте дисперсия қисығының сынуы байқалады. Яғни мынандай коэффициенттер есептеледі
Ал кейбір зерттеулер мен есептеулерде максимал электрөткізгіштік үлкен жиліктерде пайда болатындықтан, көбінесе – 104 Гц жиліктегі кедергі шамасының – шексіз жиліктегі кедергісіне қатынасын алады. Биологиялық ткань жансызданған кезде бұл көрсетілген коэффициенттер 1-ге тең болады. Қалыпты, бүлінбеген клетка мен ткандерде бұл коэффициенттің мәні эволюциялық қатардағы организмнің орнына байланысты болады. Мысалы: Сүт қоректенушілер (Мелкопитающих) - бауырларының коэффициенті К = 9 –10, құрбақаның бауырларының коэффициенті К = 2 – 3. К-коэффициентінің мәні тканнің зат алмасу қабілетіне де байланысты болады, қалыпты, бүлінбеген клетка мен ткандерде бұл коэффициенттің мәні эволюцияллық қатардағы организмнің орнына байланысты болады. Интенсивті зат алмасу органдардың (бауыр, селезенка, өт) бұл коэффициент жоғары болады, ал ол организімнің бұлшық еттерінде бұл көрсеткіш аз болады. Клетка мен ткандердің электрөткізгіштігінің дисперсиясының өзгеруі олардың поляризациялық қабілеттілігімен түсіндіріледі, сондықтан бұл жердегі келтірілген коэффициентті поляризациялық коэффициент деп атау келісілген.
|
 (1)
(1) . (2)
. (2) (3)
(3) (4)
(4) JdT = q – (5) - Бұл Т – уақыт ішінде жинақталған заряд мөлшері.
JdT = q – (5) - Бұл Т – уақыт ішінде жинақталған заряд мөлшері.

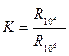 ;
;  ;
; 



