
МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ МЛЕКОПИТАЮЩИХ
К классу принадлежат наиболее высоко организованные представители подтипа позвоночных. Размеры млекопитающих изменчивы: длина тела колеблется от 3,5 см у карликовой бурозубки до 33 м у голубого кита, а вес соответственно от 1,5 г до 120 т. Тело большинства млекопитающих подразделяется на голову, шею, удлиненное туловище, опирающееся на четыре конечности и хвост (рис.1 и 2).
Рис 2 - Основные части тела млекопитающего: 1 - голова; 2 - шея; 3 - туловище; 4 - передняя конечность; 5 - задняя конечность; 6 - грудь; 7 - брюхо; 8 -пах; 9 - бок тела; 10 - спина; 11 - загривок; 12- крестец; 13 - лопатка; 14 - плечо; 15 - предплечье; 16 - запястье; 17- кисть; 18 - бедро; 19 - голень; 20 - стопа
Форма и соотношение этих частей варьирует у разных видов, отражая приспособления к среде обитания и преобладающему характеру движений, связанных с поисками и добыванием пищи, защитой от врагов и другими формами жизнедеятельности. У некоторых млекопитающих отсутствует шейный отдел (китообразные), хвост (некоторые приматы, рукокрылые) или задние конечности (китообразные, сирены). Представители разных отрядов, живущие в сходных условиях и выработавшие близкий образ жизни, могут обладать и сходной формой тела. Такие группы видов называют «жизненными формами» или экологическими типами. Имеются наземные, подземные, древесные, полуводные, околоводные, водные и летающие формы (рис.3). Так, у околоводных млекопитающих (утконос, выхухоль, бобр, ондатра, нутрия, выдра, норка) развивается густой, противостоящий намоканию мех, укорачивается шея, развиваются плавательные перепонки между пальцами, более или менее уплощается хвост. У полуводных ластоногих, особенно у водных сирен и у китообразных, формируется обтекаемое тело, конечности превращаются в ласты; развивающийся у сирен и китообразных кожистый хвостовой плавник придает им рыбообразный вид.
Рис 3 - Форма тела и характер передвижения у некоторых млекопитающих: Бег: 1 - малый суслик; 2 - заяц-русак; 3 – джейран. Лазанье по деревьям: 4 - кускус (сумчатые); 5 – белка; 6 - паукообразная обезьяна; 7 - трехпалый ленивец. Прыжки в кронах: 8 – белка; 9 – летяга, 10 - сумчатая летяга; 11 – шерстокрыл; 12 – гиббон. Полет рукокрылых: 13 - нетопырь карлик; 14 - широкоухий складчатогуб. Плавание: 15 – выхухоль; 16 - самка калана с детенышем; 17 - обыкновенный тюлень; 18 – сивуч; 19 – белуха. Подземные млекопитающие: 20 – крот; 21 – слепыш; 22 – цокор; 23 - африканский голый землекоп (по Н.Наумову, Н. Карташеву, 1979)
У наземных непарнокопытных и парнокопытных, совершающих значительные перемещения и имеющих одинаковых врагов - крупных подвижных хищников, сходный облик: высокие стройные фалангоходящие ноги, плотное туловище, длинная подвижная шея. Похожи по форме тела зайцеобразные и грызуны. Приспособление к быстрому бегу прыжками на задних конечностях (рикошетирующий бег) в открытых местообитаниях привело к выработке сходной формы тела: слабым передним и мощным задним конечностям, длинному хвосту-балансиру - у кенгуру (сумчатые), африканских прыгунчиков (насекомоядные) и различных грызунов (тушканчики, песчанки, африканские долгоноги). У роющих видов тело приобретает вальковидную форму, конечности и хвост укорачиваются (суслики, сурки и др.). При переходе к подземному образу жизни усиливаются передние конечности, участвующие в рытье (кроты, цокоры и др.). У древесных лазающих видов короткие, но сильные, снабженные острыми когтями конечности, удлиненный опушенный хвост, увеличивающий поверхность при прыжках (белки, куницы и др.). Длинные хватательные конечности приматов обеспечивают лазанье и прыжки в кронах. У опоссумов, части обезьян, древесных муравьедов, ящеров и дикобразов используется для хватания и хвост. Единственными летающими млекопитающими являются летучие мыши. У рукокрылых конечности модифицированы в крылья для полета в воздухе. У сумчатых летучих белок, летяг, шерстокрыла кожистая складка по бокам тела позволяет совершать длинные планирующие прыжки. Череп млекопитающих имеет два затылочных мыщелка, посредством которых он сочленяется с первым позвонком, что ограничивает подвижность головы (рис.4). Квадратная и сочленовная кости включаются в качестве слуховых косточек наковальни и молоточка в среднее ухо. Нижняя челюсть представлена одной зубной костью и соединяется с черепом при помощи выпуклого мыщелка, входящего в сочленовную впадину височной кости. Зубы (рис. 5) у большинства млекопитающих дифференцированы по форме и функции и разделяются, как правило, на резцы (incisivi), клыки (canini) и щечные зубы, причем последние делятся на предкоренные (premolares) и коренные (molares). Зубы располагаются в альвеолах и у многих представителей млекопитающих имеют две смены: молочную и постоянную. Число зубов, их форма и функция существенно различаются у разных групп зверей. Так, мало специализированные насекомоядные обладают большим числом сравнительно слабо дифференцированных зубов. Для грызунов и зайцеобразных характерно сильное развитие одной пары резцов (резцы постоянно растущие и не имеют корней), отсутствие клыков и плоская жевательная поверхность коренных зубов (у полевок некоторых других видов коренные зубы также не имеют корней), чтообусловлено характером питания: резцами они сгрызают растительность, а коренными зубами перетирают пищу, как жерновами. Хищные характеризуются сильно развитыми клыками, служащими для схватывания и для умерщвления добычи. Коренные зубы хищников имеют режущие вершины и плоские жевательные выступы. Задний ложнокоренной зуб верхней челюсти и первый истинно коренной нижней челюсти у хищных обычно выделяются своей величиной; они носят название плотоядных зубов.
Рис 4 - Череп (вид сверху, снизу, сбоку) и нижняя челюсть (вид сбоку) волка (вверху) и оленя благородного (внизу). Обозначения: Р - рострум, МК - мозговая капсула, Г - глазница, СД - скуловая дуга, СГ - стреловидный гребень, НВ - носовая вырезка, ЗНВ - задненёбная вырезка, ЗКМ - заглазничный костный мостик, ПГО - подглазничное отверстие, ЗРО - зарезцовое отверстие, ЭО - этмоидное отверстие, Ре - резцы, Кл - клыки, ПК - предкоренные, К - коренные, Д - диастема, ЗМ - затылочный мыщелок, ВО - венечный отросток, КО - крыловидный отросток, СО - сочленовный отросток, УО - угловой отросток (по Павлинову и др., 2002)
г - лисица), складчатая (д - лошадь, е - пищуха), лунчатая (ж - олень), призматическая (з - полевка) (по Н.Наумову, Н. Карташеву, 1979)
Общее число зубов и их распределение по группам для видов зверей вполне определенно и постоянно, служит важным признаком в систематике млекопитающих. Для упрощения описания зубов пользуются зубными формулами. Числитель показывает число зубов в верхней челюсти, знаменатель - в нижней. Группы зубов обозначаются начальными буквами их латинских названий: резцы - i, клыки – с, ложнокоренные - рm, коренные - m. Для сокращения пишется число зубов в одной половине челюсти. Зубная формула лисицы такова: i -3/3; с -1/1; pm -4/4; m -2/3= 42. Скелет. Позвонки имеют плоские сочленовные поверхности (платицельный тип позвонков). Шейный отдел позвоночника у большинства видов содержит 7 позвонков (иногда 6-9), грудной – от 12 до 15, поясничных позвонков от 2 до 9, крестцовых – 4 (рис.6). Количество хвостовых позвонков сильно варьирует.
Рис 6 - Скелет млекопитающего: 1 — череп; 2 — нижняя челюсть; 3 — шейные позвонки; 4 — грудные позвонки; 5 — поясничные позвонки; 6 — крестец; 7 — хвостовые позвонки; 8 — ребра; 9 — грудина; 10 — лопатка; 11 —плечевая кость;.12 — локтевая кость; 13 —лучевая кость; 14 — кости запястья; 15 — кости пясти; 16 — фаланги пальцев передней конечности; 17 — таз; 18 — бедренная кость; 19 — большая берцовая кость; 20 — малая берцовая кость; 21 — кости заплюсны; 22 — кости плюсны; 23 — фаланги пальцев задней конечности; 24 — коленная чашечка
У большинства млекопитающих имеется ключица, у некоторых видов она отсутствует или рудиментарна. В зависимости от того, на какой отдел конечности опираются млекопитающие, их делят на стопоходящих (медведи), пальцестопоходящих (кролики), пальцеходящих (большинство млекопитающих) и фалангоходящих (копытные). Органы чувств. Обонятельные органы у млекопитающих высоко развиты. При помощи этих органов млекопитающие опознают врагов, отыскивают пищу, а также и друг друга. Многие виды чувствуют запахи за несколько сотен метров и способны обнаруживать пищевые объекты, находящиеся под землей. Только у полностью водных зверей (китов) обоняние редуцировано. Тюлени же обладают очень острым обонянием. Прогрессивное развитие описываемых органов выражается в увеличении объема обонятельной капсулы и в ее усложнении путем образования системы обонятельных раковин. У некоторых групп зверей (сумчатые, грызуны, копытные) имеется обособленный отдел обонятельной капсулы, открывающийся самостоятельно в небно-носовой канал - так называемый якобсонов орган, который служит для восприятия вкуса пищи, находящейся во рту. В клюве утконоса имеется своеобразный «электроискатель», благодаря которому животные отыскивают под водой пищу и ориентируются в пространстве, при этом глаза и уши в водной среде у зверьков замыкаются. Аналогичный орган на голове имеется и у ехидны. Орган слуха в подавляющем большинстве случаев достигает высокого совершенства. В состав его, кроме внутреннего и среднего уха, имеющихся и у нижестоящих классов, входят еще два новых отдела: наружный слуховой проход и ушная раковина. Последняя отсутствует только у водных и подземных зверей (киты, большинство ластоногих, слепыши и некоторые другие). Ушная раковина существенно усиливает тонкость слуха. Особенно сильно она развита у ночных зверей (летучие мыши) и у видов лесных копытных, пустынных видов собачьих и некоторых других. У многих видов копытных и хищников основание наружного уха имеет трубкообразное строение, позволяющее поворачивать ушную раковину до 180º. У ряда видов зверей обнаружена способность к эхолокации. Кроме хорошо известных в этом отношении летучих мышей, такой способностью обладают китообразные (дельфины), ластоногие (тюлени), землеройки и, видимо, некоторые виды мышевидных грызунов. Землеройки при эхолокации издают импульсы частотой 30-60 кгц, продолжительностью 5-33 мсек. Органы зрения у млекопитающих не имеют существенных особенностей. Глаз у большинства млекопитающих защищен двумя непрозрачными веками (верхнее и нижнее), и, кроме того, у многих имеется прозрачная мигательная перепонка. В переднем углу глаза имеется гардерова железа, выделяющая жирный смазывающий секрет, здесь же находится и слезная железа. Зрение развито сравнительно слабо даже у обитателей открытых пространств. На неподвижные предметы звери обычно обращают мало внимания, и к стоящему человеку часто вплотную подходят такие осторожные звери, как лисицы, зайцы, лоси. Острота зрения и развитость глаз, естественно, различны и связаны с условиями существования. Особо большие глаза имеют ночные звери и виды, живущие в открытых ландшафтах (например, антилопы). У лесных видов зрение менее острое. У подземных же видов глаза редуцированы, а иногда затянуты кожистой перепонкой (слепыш, кавказский крот). Аккомодация достигается только путем изменения формы хрусталика под действием ресничной мышцы. Почти все млекопитающие хорошо видят в темноте. Для повышения светочувствительности глаза кроме «диафрагмы» - зрачка - служит еще и слой зеркального пигмента (тепетум), расположенный позади сетчатки глаза. Отражаясь от этого «зеркала», свет проходит сквозь сетчатку дважды, что вдвое улучшает способность видеть в темноте. Вот почему у многих зверей глаза ночью светятся, если на них падает луч фонаря или луны. Цветное зрение сравнительно с птицами у млекопитающих развито слабо. Почти весь спектр способны различать лишь высшие обезьяны восточного полушария. Европейская рыжая полевка различает только красный и желтый цвета. У опоссума, лесного хорька и ряда других видов цветное зрение вообще не обнаружено. Органы осязания. Характерной особенностью этих органов у млекопитающих является наличие осязательных волос. Таковыми являются вибриссы на конце морды зверей (рис. 7), а также чувствительные волосы на туловище и лапах. Кожный покров у млекопитающих имеет более сложное строение (рис. 7), чем у других позвоночных. Сложно и разнообразно и его значение. Кожный покров имеет в основном теплозащитную функцию. Волосяной покров, система кожных кровеносных сосудов, а у водных видов (киты, тюлени) - подкожный слой жира существенно предохраняют тело от излишней потери тепла. Диаметр просветов сосудов может меняться в очень больших пределах. При расширении сосудов кожи теплоотдача резко увеличивается, при сужении их теплоотдача сильно сокращается. Важное значение для охлаждения организма имеет также испарение с поверхности кожи воды (пота), выделенного потовыми железами.
1 - наружный слой рогового эпидермиса, клеточки которого спадают время от времени; 2 - глубокий слой эпидермиса с живыми клетками (мальпигиев слой); 3 -собственно кожа (кориум); 4 - волос; 5 - потовая железа; 6 - отверстие ее протока; 7 - сальная железа; 9 - мышцы волоса; 9 - соединительнотканые волокна кожи; 10 - кровеносный сосуд; 11 - сосочек в основании волоса (по С. Наумову, 1973)
Температура тела у многих млекопитающих относительно постоянна, и ее отличие от температуры внешней среды может достигать 100°C. Так, у песца, обитающего зимой при температуре до минус 65°С, температура тела равна +36-39°С. Постоянство температуры тела (гомотермия) не является абсолютным признаком всех млекопитающих. В полной мере она характерна для видов, имеющих относительно крупные размеры. У низших млекопитающих менее развит терморегуляционный механизм, а у мелких плацентарных зверей, имеющих невыгодное для сохранения тепла соотношение между объемом тела и его поверхностью, температура тела меняется в значительных пределах в зависимости от температуры внешней среды. Так, у сумчатой крысы температура тела меняется в пределах 37,8-29,3°С, у наиболее примитивных насекомоядных (тенреков) - 34-13°С, у одного из видов броненосцев - 40-27°С, у обыкновенной полевки - 37-32°С. Как и у других позвоночных, кожа млекопитающих состоит из двух слоев: наружного (эпидермис) и внутреннего (кориум, или собственно кожа). Поверхностно расположенные клетки перерождаются, ороговевают и постепенно снашиваются в виде мелкой «перхоти» или целыми лоскутами (так, например, бывает у тюленей). Снашивание рогового слоя эпидермиса восполняется постоянным нарастанием его за счет деления клеток мальпигиева слоя. Эпидермис дает начало многим производным кожи, основные из которых - волосы, ногти, когти, копыта, рога (кроме оленьих), чешуи, различные железы. Собственно кожа, или кориум, у млекопитающих развита очень сильно. Она состоит преимущественно из волокнистой соединительной ткани, сплетения волокон которой образуют сложный узор. Нижняя часть кориума состоит из очень рыхлой волокнистой ткани, в которой откладывается жир. Слой этот носит название подкожной жировой клетчатки. Наибольшего развития он достигает у водных зверей - китов, тюленей, у которых в связи с полной (у китов) или частичной (у тюленей) редукцией волосяного покрова выполняет термоизоляционную роль. У некоторых наземных зверей также бывают большие подкожные жировые отложения. Особенно сильно они развиты у видов, впадающих на зиму в спячку или сон (суслики, сурки, медведи, барсуки и др.). Для них жир во время спячки служит основным энергетическим и питательным материалом. Толщина кожи различна у разных видов. Как правило, у видов холодных стран, имеющих пышный волосяной покров, она тоньше. Очень тонкая и непрочная кожа характерна для зайцев, к тому же она бедна кровеносными сосудами. Это имеет приспособительное значение, выражающееся в своеобразной автотомии. Хищник, схватив зайца за кожу, легко вырывает из нее кусок, упуская самого зверька. Образовавшаяся рана почти не кровоточит и быстро заживает. Своеобразная кожная хвостовая автотомия наблюдается у некоторых мышей, сонь, тушканчиков. Кожный хвостовой футляр у них легко обрывается и соскальзывает с хвостовых позвонков, что позволяет схваченному за хвост зверьку уйти от врага. Волосяной покров характерен для большинства млекопитающих. Лишь немногие виды вторично, полностью или частично утратили волосяной покров. Совсем не имеют волос дельфины, у китов есть только зачатки волос на губах. У ластоногих волосяной покров редуцированный, что отмечается у моржей, в наименьшей мере - у ушастых тюленей (например, у котика), которые более, чем другие виды ластоногих, связаны с сушей. По длине волоса можно различить ствол - часть, выступающую над кожей, и корень - часть, сидящую в коже. Нарастание волос идет за счет размножения и видоизменения клеток луковицы в корне волоса. Ствол волоса является уже мертвым роговым образованием, не способным к росту и изменению формы. Погруженный в кожу корень волоса сидит в волосяной сумке, наружные стенки которой состоят из наружного слоя, или волосяного мешка. В волосяной мешок открываются протоки сальных желез, секрет которых смазывает волос и сообщает ему большую прочность и водоупорность. К нижней части волосяного мешка прикрепляются мышечные волокна, сокращения которых способствуют движению мешка и сидящего в нем волоса. Это движение и обусловливает «ощетинивание» зверя. Обычно вершины волос направлены в какую-либо сторону, так что волосы более или менее прилегают к коже. Неравномерность в расположении волос выражена не у всех видов одинаково. Наименее заметна она у подземных зверей, например, у крота. Волосяной покров состоит из различных типов волос. Наиболее толстые и длинные - это кроющие: направляющие и остевые имеют «копьевидную» форму. Они поддерживают самые многочисленные, тонкие и извитые пуховые волосы. В воде верхняя часть остевых волос отклоняется в противоположную движению животного сторону и ложится на пружинящий слой пуховых волос и предохраняет их от намокания и механических повреждений (рис. 8).
Рис 8 - Положение волос у полуводных зверей: а – на суше; б – при плавании и нырянии: 1- направляющие и остевые, 2- пуховые волосы (по Н.Наумову, Н. Карташеву, 1979)
У подземных зверей (крот, слепыш) меховой покров почти полностью лишен остевых волос. Наоборот, у взрослых оленей, кабанов и тюленей редуцирован подшерсток и покров состоит главным образом из остей. У молодых особей этих зверей подшерсток развит хорошо. Шерсть млекопитающих - совершенное изоляционное покрытие. В экстренных случаях животные резко повышают выработку тепла с помощью работы мышц - мускульной дрожи, а также взъерошивают волосяной покров, делая теплоизолирующий слой шерсти более толстым и рыхлым. Такой же прием срабатывает при сильном испуге: чтобы показаться большим и страшным противнику, шерсть или волосы встают дыбом. Волосяной покров периодически меняется. По многим видам нет ясности о числе линек. Животные с сезонным диморфизмом окраски и полным побелением волосяного покрова - песец, горностай, ласка - имеют две линьки (весенняя и осенняя), но некоторые авторы считают, что у зайца-беляка осенью не происходит выпадение летнего меха, а идет его побеление и доращивание дополнительных пуховых волос. То есть у зайцев и пищух одна линька. Животные без изменения пигментации - куницы, соболь, колонок, солонгой, перевязка, хорьки, харза, росомаха, все кошачьи, волк, шакал, гиена, корсак, тонкопалый суслик, слепыши, цокоры, кроты - имеют по две линьки. Но и по этим видам много разночтений. Доказано, что у лисицы одна полная линька, а осенью удлиняется летний волос и дополнительно вырастают пуховые волосы. Сомнения относительно двух линек высказываются по волку, шакалу, гиене, солонгою, слепышам и цокору. У кротов отмечается третья компенсационная линька. У всех зимоспящих одна полная линька. У медведей в берлоге происходит полное слущивание эпидермиса с подошв лап, а летом – смена волос. На сурках доказано, что происходит одна полная смена волос со слущиванием эпидермиса летом, а в спячке – только слущивание эпидермиса на всем теле и на подошвах лап. Аналогичные процессы возможны и у других зимоспящих. Полуводные и ластоногие имеют одну растянутую малозаметную смену волос. По копытным нет единого мнения. Одни исследователи считают, что у них одна полная линька с доращиванием волос осенью, а другие – что две полные линьки. У благородного оленя две линьки на туловище и одна на голове и конечностях.
Особую категорию представляют чувствующие волосы, или вибриссы, - длинные жесткие волосы, выполняющие осязательную функцию (рис. 9).
Рис 9 - Вибриссы на морде зверя образуют «осязательную зону» (по С.Наумову, 1973)
Такие волосы расположены чаще на голове («усы»), на нижней части шеи, на груди, а у некоторых лазающих древесных форм (например, у белки) и на брюхе. В основании волосяного мешка в его стенках располагаются мякотные нервные волокна, воспринимающие соприкосновения стержня вибриссы с посторонними предметами. Видоизменениями волос являются щетина и иглы. Прочие производные эпидермиса представлены чешуей, ногтями, когтями, копытами, «полыми» рогами, роговым клювом. Чешуя зверей по своему развитию и строению сходна с одноименным образованием рептилий. Наиболее сильно развита чешуя у ящеров, у которых она покрывает все тело. У многих мышевидных чешуйки имеются на лапах. Наличие чешуи на хвосте характерно для многих сумчатых, грызунов и насекомоядных. Роговыми образованиями являются также «полые» рога быков, антилоп, козлов и баранов. Они развиваются из эпидермиса и сидят на костных стержнях, представляющих самостоятельные кости, сросшиеся с лобными костями. Рога оленей имеют иную природу. Они развиваются из кориума и состоят из костного вещества. Кожные железы у млекопитающих весьма многочисленны и разнообразны по строению и функции. Основные типы желез следующие: потовые, сальные, пахучие, млечные. Потовые железы трубчатые, глубинные части их имеют вид клубка. Они открываются непосредственно на поверхности кожи или в волосяную сумку (рис. 5). Продуктом выделения этих желез является пот, состоящий в основном из воды, в которой растворены мочевина и соли. Эти продукты не вырабатываются клетками желез, а поступают в них из кровеносных сосудов. Функция потовых желез заключается в охлаждении тела путем испарения выделяемой ими на поверхность кожи воды и в выделении продуктов распада. Эти железы совмещают в себе терморегуляционную и выделительную функции. Потовые железы есть у большинства млекопитающих, но развиты они не у всех одинаково. Их очень мало у собак и кошек; у многих грызунов они имеются только на подошвах лап, в паховой области и на губах. Отсутствуют потовые железы у китообразных, ящеров и некоторых других. Сальные железы имеют гроздевидное строение и открываются в волосяную сумку (рис. 5). Жирный секрет желез смазывает волос и верхний слой эпидермиса кожи, предохраняя их от смачивания и снашивания. Пахучие железы представляют видоизменение потовых или сальных желез, а иногда комбинацию тех и других. Например, анальные железы куньих, секрет которых имеет очень резкий запах и служит в основном для защиты от преследующих врагов. Особенно сильно развиты эти железы у американских скунсов, способных выпрыскивать большие порции выделений на значительное расстояние. Мускусные железы имеются у кабарги, выхухоли, бобра, ондатры и др. Судя по тому, что наибольшее развитие они получают в период гона, деятельность их, видимо, связана с размножением; возможно, они стимулируют половое возбуждение и привлекают особей противоположного пола. Пахучие секреты желез используются для самомаркировки зверей, мечения границ и территории обитания, обеспечения хемокоммуникаций биологического «сигнального поля». Млечные железы представляют своеобразное видоизменение простых трубчатых потовых желез. В простейшем случае - у австралийских однопроходных - они сохраняют трубчатое строение и открываются в сумки волос, расположенных группами на небольшом участке брюшной поверхности, так называемом железистом поле. Сосков у однопроходных нет, и молоко слизывается детенышами с волос, куда оно поступает из волосяных сумок. У сумчатых и плацентарных млечные железы имеют гроздевидное строение и протоки их открываются на сосках. Расположение желез и сосков бывает различным. У лазающих по деревьям обезьян и висящих во время кормления летучих мышей имеется только пара сосков на груди, у бегающих копытных соски располагаются только в паховой области. У насекомоядных и хищных соски располагаются двумя рядами по всей нижней поверхности туловища. Число сосков стоит в прямой связи с плодовитостью вида и в некоторой степени соответствует числу одновременно рождающихся детенышей. Минимальное число сосков (два) у обезьян, овец, коз, слонов и некоторых других; максимальное число сосков (10-24) у мышевидных грызунов, насекомоядных, некоторых сумчатых. Органы пищеварения характеризуются общим удлинением пищеварительного тракта, большей, чем у других позвоночных, его дифференцировкой и большей развитостью пищеварительных желез. Пищеварительный тракт начинается преддверием рта, расположенным между свойственными только млекопитающим мясистыми губами, щеками и челюстями. У ряда видов преддверие, расширяясь, образует большие защечные мешки (хомяки, бурундуки, обезьяны). Мясистые губы служат для схватывания пищи, а преддверие рта для временного ее резервирования; так, хомяки и бурундуки переносят в защечных мешках запасы пищи в свои норы. Мясистых губ нет у однопроходных и китообразных. В ротовой полости пища подвергается механическому измельчению и химическому воздействию. У зверей имеются четыре пары слюнных желез, секрет которых содержит фермент птиалин, превращающий крахмал в декстрин и мальтозу. Развитость слюнных желез зависит от характера питания. У китообразных они практически не развиты; у жвачных же, наоборот, они получили исключительно сильное развитие. Так, корова в сутки выделяет около 56 л слюны, которая имеет огромное значение для смачивания грубой пищи и для наполнения жидкой средой полостей желудка, где происходит бактериальное расщепление клетчатки пищевой массы. Секрет щечных желез летучих мышей, наносимый на летательные перепонки, сохраняет их эластичность и предохраняет от высыхания. Слюна вампиров, питающихся кровью, обладает антикоагулирующими свойствами, т.е. предохраняет кровь от свертывания. Слюна некоторых землероек ядовита; секрет их подчелюстной железы вызывает смерть мыши менее чем через 1 минуту после инъекции. Токсичность слюнных желез примитивных млекопитающих рассматривается как отражение их филогенетической связи с рептилиями. Желудок обособлен от других отделов пищеварительного тракта и снабжен многочисленными железами. Объем желудка и его внутреннее строение различны у разных видов, что связано с характером пищи. Наиболее просто устроен желудок у однопроходных, имеющий вид простого мешка. У большинства желудок разделен на большее или меньшее число отделов. Усложнение желудка связано со специализацией питания, например, с поглощением огромной массы грубых кормов (жвачные) или недоразвитостью ротового пережевывания пищи (некоторые виды, питающиеся насекомыми). У некоторых южноамериканских муравьедов в выходной части желудка дифференцируется отдел с настолько твердыми складками, что они выполняют функцию зубов, перетирающих пищу. Собственно кишечник подразделяется на тонкий, толстый и прямой отделы. У видов, питающихся грубым растительным кормом (например, у грызунов), от границы тонкого и толстого отделов отходит длинная и широкая слепая кишка, заканчивающаяся у некоторых зверей (например, зайцев, полуобезьян) червеобразным отростком. Слепая кишка выполняет роль бродильного чана и развита тем сильнее, чем больше растительной клетчатки поглощает при кормлении животное. У видов мышей, питающихся семенами и отчасти вегетативными частями растений, слепая кишка составляет 7-10% от общей длины всех отделов кишечника, а у полевок, кормящихся в большей мере вегетативными частями растений, - 18-27%. У плотоядных видов слепая кишка развита слабо или отсутствует. Варьирует и длина толстого отдела кишечника. У грызунов он составляет 29-52% общей длины кишечного тракта, у насекомоядных и рукокрылых - 26-30%, у хищных - 13-22%. Общая длина кишечника варьирует очень сильно. У растительноядных видов кишечник относительно более длинный, чем у всеядных и плотоядных видов. Так, у некоторых летучих мышей кишечник длиннее тела в 2,5 раза, у насекомоядных - в 2,5-4,2 раза, у хищных - в 2,5 раза (ласка), в 6,3 раза (собака), у грызунов - в 5 раз (полуденная песчанка), в 11,5 раз (морская свинка), у лошади - в 12 раз, у овцы - в 29 раз. Выделительная система. Почки у млекопитающих тазовые. Основным конечным продуктом белкового обмена у млекопитающих (как и у рыб и амфибий) в отличие от рептилий и птиц бывает не мочевая кислота, а мочевина. Если у птиц долевое значение мочевой кислоты равно 63-80% и на долю мочевины приходится 1-10%, то у млекопитающих соотношение обратное: мочевина - 68-91%, мочевая кислота - 0,1-8%. Выделительную функцию частично выполняют и потовые железы, через которые выводятся растворы солей и мочевина. Этим путем выводится не более 3% азотистых продуктов белкового обмена. Органы воспроизведения. Половые железы самца - семенники - имеют характерную овальную форму. У однопроходных, некоторых насекомоядных и неполнозубых, у слонов и китообразных они в течение всей жизни находятся в полости тела. У большинства других зверей семенники первоначально располагаются в полости тела, но по мере полового созревания они опускаются вниз и попадают в особый наружно расположенный мешочек - мошонку (scrotum). У многих млекопитающих прочность полового члена обусловливается еще особой длинной костью (os penis), расположенной между пещеристыми телами. Таковы хищные, ластоногие, многие грызуны, некоторые летучие мыши и др. Парные яичники всегда лежат в полости тела и прикреплены к спинной стороне брюшной полости брыжейками. Верхний отдел яйцеводов представляет фаллопиевы трубы. Далее идут расширенные отделы - матки, которые открываются в непарный у большинства зверей отдел - влагалище. Последнее переходит в короткий мочеполовой канал, в который, кроме него, открывается мочеиспускательный канал. На брюшной стороне мочеполового канала располагается небольшой вырост - клитор (clitoris), обладающий пещеристыми телами и соответствующий пенису самца. Любопытно, что у некоторых видов в клиторе имеется кость (os с1itoridis). Плацента. Во время развития эмбриона в матке млекопитающих формируется образование, называемое плацента. Только у однопроходных плацента отсутствует. У сумчатых есть зачатки плаценты. Плацента возникает путем срастания наружной стенки аллантоиса с серозой, в результате чего формируется губчатое образование - хорион. Хорион образует выросты - ворсинки, которые соединяются или срастаются с разрыхленным участком эпителия матки. В этих местах кровеносные сосуды детского и материнского организмов сплетаются (но не сливаются), и таким образом устанавливается связь между кровяными руслами эмбриона и родителя. В результате этого обеспечивается газообмен в теле зародыша, его питание и удаление продуктов распада. Плацента свойственна уже сумчатым млекопитающим, хотя у них она еще примитивная. У высших плацентарных зверей хорион всегда образует выросты - ворсинки, соединяющиеся со стенками матки. Выделяют несколько типов плаценты: диффузная, когда ворсинки распределяются равномерно по хориону (китообразные, многие копытные, полуобезьяны); дольчатая, когда ворсинки собраны в группы, распределенные по всей поверхности хориона (большинство жвачных); дискоидальная, когда ворсинки располагаются на ограниченном, имеющем вид диска участке хориона (насекомоядные, грызуны, обезьяны). Степень связанности детской и материнской плацент различна. В одних случаях эта связь непрочная, и при родах слизистая матки не отрывается. В других - связь более прочная, и при родах соответствующая часть слизистой матки отпадает и выводится наружу как послед. Первая плацента называется неотпадающей, вторая - отпадающей. Строение женских половых путей различается у разных групп млекопитающих. У однопроходных яйцеводы на всем протяжении парные и дифференцированы только на фаллопиевы трубы и матки, которые самостоятельными отверстиями открываются в мочеполовой синус. У сумчатых обособляется влагалище, но часто оно остается парным. У плацентарных влагалище всегда непарное, а более верхние отделы яйцеводов в той или иной мере сохраняют парный характер. В простейшем случае матка парная, и левый и правый ее отделы открываются во влагалище самостоятельными отверстиями. Такая матка называется двойной; она свойственна многим грызунам, некоторым неполнозубым. Матки могут быть соединены только в нижнем отделе - двураздельная матка. Подобное явление наблюдается у некоторых грызунов, летучих мышей, хищников. Слияние значительной части левой и правой маток приводит к образованию двурогой матки хищных, китообразных, копытных. Наконец, у приматов, полуобезьян и некоторых летучих мышей матка непарная - простая, парными остаются только верхние отделы яйцеводов - фаллопиевы трубы.
СХЕМА ИЗМЕРЕНИЙ ЧАСТЕЙ ТЕЛА НАЗЕМНЫХ МЛЕКОПИТАЮЩИХ
1. Длина тела - от верхнего края губы до корня хвоста (по изгибам тела мерной лентой). У видов по размерам меньше зайца – со стороны живота мерной линейкой или штангенциркулем. 2. Длина хвоста – от основания хвоста до его кончика без концевых волос. 3. Длина ступни - от пяточного бугра до кончика самого длинного пальца (копыта) без когтя (мерной линейкой или штангенциркулем). 4. Длина уха – от нижнего края ушной вырезки до кончика без волос. 5. Высота в холке – от хребта над лопаткой до конца копыта или самого длинного пальца без когтей. Нога вытягивается перпендикулярно продольной оси тела (мерной лентой или линейкой). 6. Охват туловища – позади основания передних конечностей (мерной лентой). 7. Косая длина туловища – от переднего края плечелопаточного сустава до заднего выступа седалищного бугра (мерной лентой). 8. Длина передней ноги - в вытянутом состоянии от вершины локтевого сустава до кончика копыта. 9. Длина задней ноги – в естественном состоянии от выступа тазобедренного сустава до кончика копыта (у копытных) самого длинного пальца через коленный и пяточный суставы. 10. Длина кисти – от плюсневого бугра до кончика копыта (мерной лентой или линейкой).
После овуляции на месте лопнувшего фолликула образуется желтое тело овуляции. Если происходит оплодотворение яйцеклетки, то желтое тело овуляции превращается в желтое тело беременности. У некоторых млекопитающих яйцеклетка после оплодотворения задерживается в развитии и не имплантируется в слизистую оболочку матки. Наступает диапауза (латентный период), продолжительность которой у разных видов неодинакова. После окончания диапаузы происходит имплантация бластоциты в стенку матки и ее нормальное развитие.
СХЕМА ПРОМЕРОВ ТЕЛА ЛАСТОНОГИХ
Рис 11 - Вид сбоку (по Млекопитающие фауны СССР, 1963, ч.2): 1— длина тела от конца морды (носа) до конца хвоста по спинной поверхности тела; 2 — длина тела между теми же точками по горизонтальной плоскости; 3— длина тела от конца морды (носа) до конца задних ластов по спинной поверхности тела; 4— поперечный (подмышечный) обхват тела; 5— наибольший обхват тела; 6 — длина переднего ласта по переднему краю; 7— длина переднего ласта от подмышечной впадины; 8 — длина заднего ласта (от его основания); 9— длина хвоста (от его основания)
|

 Рис 1 - Сравнительные размеры животных, в том числе и разных видов млекопитающих: 1- нильский крокодил; 2 - странствующий альбатрос; 3 - слоновая черепаха; 4 – человек; 5 - императорский пингвин; 6 - африканский страус; 7 – капибара; 8 - серый кенгуру; 9 - белый медведь; 10 – тигр; 11 – лось; 12 – жираф; 13 - слон африканский; 14 – анаконда
Рис 1 - Сравнительные размеры животных, в том числе и разных видов млекопитающих: 1- нильский крокодил; 2 - странствующий альбатрос; 3 - слоновая черепаха; 4 – человек; 5 - императорский пингвин; 6 - африканский страус; 7 – капибара; 8 - серый кенгуру; 9 - белый медведь; 10 – тигр; 11 – лось; 12 – жираф; 13 - слон африканский; 14 – анаконда

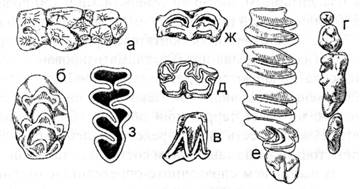 Рис 5 - Основные варианты жевательной поверхности коренных зубов у млекопитающих: бугорчатая (а - кабан, б - лесная мышь), бугорчато-режущая (в - кожан,
Рис 5 - Основные варианты жевательной поверхности коренных зубов у млекопитающих: бугорчатая (а - кабан, б - лесная мышь), бугорчато-режущая (в - кожан,
 Рис 7 - Разрез кожи млекопитающего:
Рис 7 - Разрез кожи млекопитающего: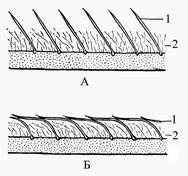
 Густота и высота волосяного покрова у животных меняется по сезонам: у белки на 1 см2 огузка летом в среднем 4200 волос, зимой - 8100, у зайца-беляка - 8000 и 14700. Длина волос в миллиметрах на огузке такова: у белки летом пух - 9,4, ость - 17,4, зимой - 16,8 и 25,9; у зайца-беляка летом пух - 12,3, ость - 26,4, зимой - 21,0 и 33,4. У тропических зверей таких резких изменений не бывает из-за малой разницы в температурных условиях зимой и летом.
Густота и высота волосяного покрова у животных меняется по сезонам: у белки на 1 см2 огузка летом в среднем 4200 волос, зимой - 8100, у зайца-беляка - 8000 и 14700. Длина волос в миллиметрах на огузке такова: у белки летом пух - 9,4, ость - 17,4, зимой - 16,8 и 25,9; у зайца-беляка летом пух - 12,3, ость - 26,4, зимой - 21,0 и 33,4. У тропических зверей таких резких изменений не бывает из-за малой разницы в температурных условиях зимой и летом.
 Рис 10 - 1- длина тела; 2 - длина хвоста; 3 - длина ступни; 4 - длина уха; 5 - высота в холке; 6 - охват туловища; 7 - косая длина туловища; 8 - длина передней ноги; 9 - длина задней ноги; 10 – длина кисти
Рис 10 - 1- длина тела; 2 - длина хвоста; 3 - длина ступни; 4 - длина уха; 5 - высота в холке; 6 - охват туловища; 7 - косая длина туловища; 8 - длина передней ноги; 9 - длина задней ноги; 10 – длина кисти




