Введение. Задание 1. Перечислите в таблице функции стебля
Задание 1. Перечислите в таблице функции стебля. Какая ткань участвуют в выполнении каждой функции?
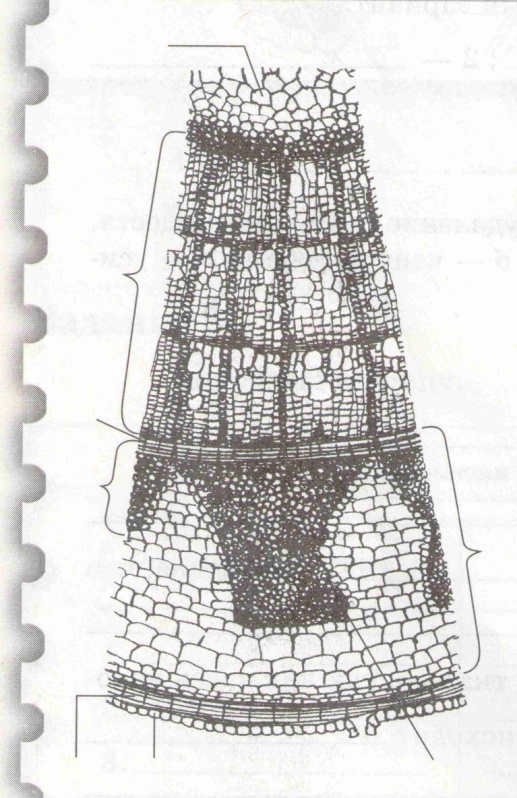
Задание 2. Рассмотрите рисунок поперечного среза стебля липы. Определите части стебля. Расставьте соответствующие цифры на рисунке.
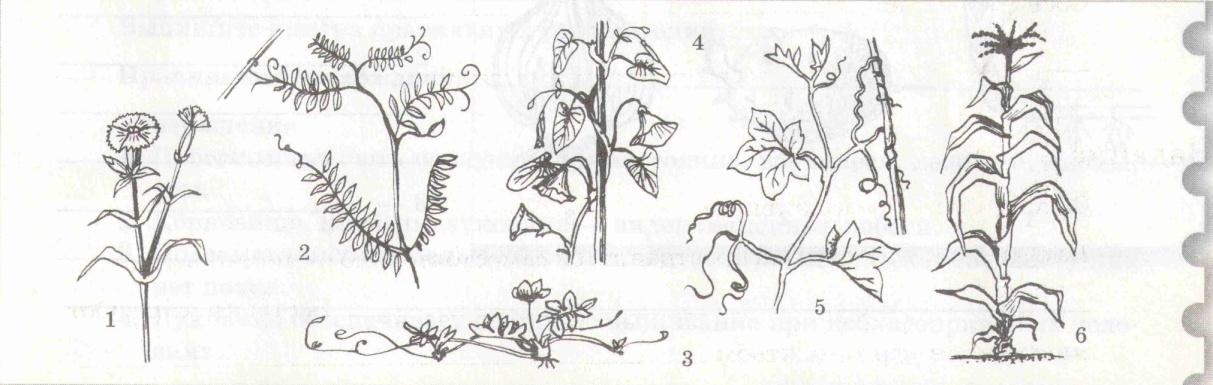
Задание 3. Напишите, какие условия жизни растений (факторы среды – биотические, абиотические) могут повлиять на изменение формы стеблей у растений. Приведите 2-3 примера. ________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________ Задание 4. Напишите названия типов стеблей, указанных на рисунке.
Введение Последние десятилетия XX века ознаменовались бурным развитием одной из главных ветвей биологической науки - молекулярной генетики. Уже в начале 70-х годов ученые в лабораторных условиях начали получать и клонировать рекомбинантные молекулы ДНК, культивировать в пробирках клетки и ткани растений и животных. Возникло новое направление генетики - генетическая инженерия. На основе ее методологии начали разрабатываться различного рода биотехнологии, создаваться генетически измененные организмы (ГМО). Появилась возможность генной терапии некоторых заболеваний человека, а последнее десятилетие XX века ознаменовалось еще одним важным событием - достигнут огромный прогресс в клонировании животных из соматических клеток. Разработанные методы клонирования животных пока еще далеко не совершенны. В процессе экспериментов наблюдается высокая смертность плодов и новорожденных. Еще не ясны многие теоретические вопросы клонирования животных из отдельной соматической клетки. Тем не менее, многие ученые с энтузиазмом восприняли идею клонирования человека. Опрос общественного мнения в США показал, что 7% американцев готовы подвергнуться клонированию. Вместе с тем, большинство ученых и многие политики высказываются против создания клонов человека. И их возражения и опасения вполне оправданы. Клон – (от греч. сlon – отпрыск, ветвь) это группа клеток или организмов, происшедших от общего предка путём бесполого размножения и являющихся генетически идентичными. Примером клона можно назвать группу бактериальных клеток, образовавшихся в результате деления исходной клетки, потомков морской звезды, регенерировавших из частей разделённого материнского организма, клоном также являются все кусты или деревья, полученные путём вегетативного размножения. Однако вот млекопитающим способность размножаться путём клонирования природа не "предусмотрела". Высокий уровень дифференциации клеток как бы "обратной стороной медали" обозначает утрату ними способности давать начало новому организму. Однако, как показала практика, ядро даже дифференцированной клетки сохраняет все потенции, необходимые для того, чтобы дать начало новому организму. Первые опыты на амфибиях Возможность клонирования эмбрионов позвоночных впервые была показана в начале 50-х годов в опытах на амфибиях. Американские исследователи Бриггс и Кинг разработали микрохирургический метод пересадки ядер эмбриональных клеток с помощью тонкой стеклянной пипетки в лишенные ядра (энуклеированные) яйцеклетки. Они установили, что если брать ядра из клеток зародыша на ранней стадии его развития - бластуле, то примерно в 80% случаев зародыш благополучно развивается дальше и превращается в нормального головастика.Если же развитие зародыша, донора ядра, продвинулось на следующую стадию - гаструлу, то лишь менее чем в 20% случаев оперированные яйцеклетки развивались нормально. Эти результаты позже были подтверждены и в других работах.Большой вклад в эту область внес английский биолог Гердон. Он первым в опытах с южноафриканскими жабами Xenopus laevis (1962) в качестве донора ядер использовал не зародышевые клетки, а уже вполне специализировавшиеся клетки эпителия кишечника плавающего головастика. Ядра яйцеклеток реципиентов он не удалял хирургическим путем, а разрушал ультрафиолетовыми лучами. В большинстве случаев реконструированные яйцеклетки не развивались, но примерно десятая часть их них образовывала эмбрионы. 6,5% из этих эмбрионов достигали стадии бластулы, 2,5% - стадии головастика и только 1% развился в половозрелых особей.Однако появление нескольких взрослых особей в таких условиях могло быть связано с тем, что среди клеток эпителия кишечника развивающегося головастика довольно длительное время присутствуют первичные половые клетки, ядра которых могли быть использованы для пересадки. В последующих работах как сам автор, так и многие другие исследователи не смогли подтвердить данные этих первых опытов.Позже Гердон модифицировал эксперимент. Поскольку большинство реконструированных яйцеклеток (с ядром клетки кишечного эпителия) погибают до завершения стадии гаструлы, он попробовал извлечь из них ядра на стадии бластулы и снова пересадить их в новые энуклеированные яйцеклетки (такая процедура называется "серийной пересадкой" в отличие от "первичной пересадки"). Число зародышей с нормальным развитием после этого увеличивалось, и они развивались до более поздних стадий по сравнению с зародышами, полученными в результате первичной пересадки ядер.Затем Гердон вместе с Ласки (1970) стали культивировать in vitro (вне организма в питательной среде) клетки почки, легкого и кожи взрослых животных и использовать уже эти клетки в качестве доноров ядер. Примерно 25% первично реконструированных яйцеклеток развивались до стадии бластулы. При серийных пересадках они развивались до стадии плавающего головастика. Таким образом, было показано, что клетки трех разных тканей взрослого позвоночного (X. laevis) содержат ядра, которые могут обеспечить развитие по крайней мере до стадии головастика.В сною очередь ДиБерардино и Хофнер использовали для трансплантации ядра недслящихся и полносгью дифференцированных клеток крови - эритроцитов лягушки Rana pipiens. После серийной пересадки таких ядер 10% реконструированных яйцеклеток достигали стадии плавающего головастика. Однако даже с помощью многократных серийных пересадок (более 100 клеточных циклов) реконструированные яйцеклетки дальше стадии головастика не развивались.Таким образом, во многих работах показано, что в случае амфибий донорами ядер могут быть лишь зародыши на ранних стадиях развития. Некоторые авторы называют подобные эксперименты клонированием амфибий, хотя правильнее называть их клонированием эмбрионов амфибий, так как в этом случае мы размножаем бесполым путем не взрослых животных, а зародышей.Дифференцировка клеток в ходе развития позвоночных сопровождается инактивацией неработающих генов. Поэтому дифференцировка становится необратимой. В конце концов у одних клеток происходит полное репрессирование генома, у других - в той или иной степени деградирует ДНК, а в некоторых случаях разрушается даже ядро. Однако наряду с дифференцированными кочетками культивируемые in vitro клеточные популяции содержат малодифференцированные стволовые клетки, которые и могут быть использованы как доноры ядер для клонирования млекопитающих.Опыты с амфибиями показали, что ядра различных типов клеток одного и того же организма генетически идентичны и в процессе клеточной дифференцировки постепенно теряют способность обеспечивать развитие реконструированных яйцеклеток, однако серийные пересадки ядер и культивирование клеток invitro в какой-то степени увеличивает эту способность. Неудачи экспериментов с мышами Успешные опыты с амфибиями заставили ученых задуматься о клонировании эмбрионов млекопитающих, в частности мышей. МакКиннел в одной из своих работ отмечал, что все необходимые для этого методы уже существуют, и непонятно, почему мышь до сих пор не клонирована. По его мнению, первыми объектами должны были стать именно мелкие животные, такие как мышь или кролик. Однако предсказание МакКиннелла не сбылось, хотя в конце 70-х годов опыты на мышах действительно начались и протекали весьма драматично. К тому времени, замечу, весьма основательно были изучены биология и генетика ранних этапов развития млекопитающих, и, в частности, мыши как модельного объекта. Работа методически оказалась довольно трудной, прежде всего потому, что объем яйцеклетки у млекопитающих примерно в тысячу раз меньше, чем у амфибий.Однако эти трудности были успешно преодолены. Экспериментаторы научились микрохирургически удалять пронуклеусы из зигот (оплодотворенных яйцеклеток) мыши и пересаживать в них клеточные ядра ранних эмбрионов. Однако все полученные разными способами зародыши мышей развивались лишь до стадии бластоцисты.Пронуклеус - одно из двух гаплоидных ядер в яйце млекопитающих в период послепроникновения сперматозоида, но до слияния мужского и женского пронуклеусов в ядро зиготы в процессе оплодотворения. Мужское ядро формируется из ядерного материала сперматозоида, женское - из хромосом яйцеклетки.Бластоциста (бластула) - зародыш млекопитающих на одной из ранних стадий развития, еще до его имплантации в матку.В 1977 году появилось сенсационное сообщение Хоппе и Илменси о том, что они получили семь взрослых самок мышей, пять из которых имели голько магеринский, а две - отцовский геном. Это, якобы, зависело от гого, какой пронуклеус был оставлен в яйце - женский или мужской, он и определял развитие особи но типу гиногенеза или андрогенеза. Их успех был связан, но описанию авторов, с гем, что, удаляя один нронуклеус, они удваивали число хромосом другого, обрабатывая яйца специальным веществом, затем выращивали полученные диплоидные гомочиготные (с двумя одинаковыми наборами генов) зародыши in vitro до стадии бластоцисты и пересаживали в матку самки-реципиента для дальнейшего развития.Казалось, теперь можно будет быстро получать млекопитающих со 100%-ной гомозиготностью по всем генам. Это особенно важно в селекции, так как для получения сельскохозяйственных животных, в частности, крупного рогатого скота, с закрепленными особо ценными качествами обычными приемами требуются десятки лет работы.Однако, к сожалению, данные Хоппе и Илменси подтвердить не удалось, хотя многие пытались это сделать. Оказалось, что полученные любым способом диплоидные андрогенетические и гиногенетические зародыши мышей погибают на тех же стадиях, что и диплоидные партеногенетические (развивающиеся из неоплодотворенной яйцеклетки) эмбрионы.Значительно усовершенствовав методы извлечения ядер и введения их в клетку, МакГрат и Солтер провели свою серию экспериментов и сообщили, что высокий выход живых мышей они получили, когда в качестве доноров ядер использовали зиготы, но если донорами были ранние эмбрионы, то реконструированные яйцеклетки, как и прежде, развивались только до стадии бластоцисты.Метод МакГрата и Солтера стал широко использоваться разными экспериментаторами. Так, Манн и Ловел-Бадж выделяли пронуклеусы из яиц, активированных к партеногенезу, и пересаживали их энуклеированные зиготы мышей. В этих случаях эмбрионы погибали на ранних стадиях. Если же наоборот, пронуклеусы получали из оплодотворенных яиц и пересаживали в партеногенетически активированные и лишенные ядра яйца, то такие зародыши развивались нормально до рождения. Сурани с соавторами установили, что если добавить женский пронуклеус из зиготы мыши к гаплоидному набору хромосом яйцеклетки, то нормального развития не происходит, добавление же мужского ядра приводит к нормальному развитию. С другой стороны, рекомбинации мужского и женского пронуклеусов из разных оплодотворенных яйцеклеток мышей обеспечивает нормальное развитие, а комбинация двух мужских или двух женских пронуклеусов останавливает развитие эмбриона.Эти опыты показали, что для нормального развития млекопитающих требуются два набора хромосом - отцовский и материнский. Поэтому ни у одного из известных видов млекопитающих не описан партеногенез. Поэтому работы Хоппе и Илменси не удалось повторить.Однако эти исследователи еще дважды будоражили научное сообщество. В 1982 году они пересадили ядра клеток партеногенетических бластоцист мышей вэнуклеированные зиготы Некоторые из этих реконструированных яйцеклеток нормально развивались, и якобы были получены четыре взрослых самки. В свете вышесказанного эти результаты весьма маловероятны.Гибель партеногенетических (гиногенетических) и андрогенетических зародышей у млекопитающих связана с различной активностью в онтогенезе материнского и отцовского геномов. Механизм, регулирующий эти функциональные различия, был назван геномным импринтингом и изучался в ряде работ, где было показано, что для нормального развития млекопитающих требуется наличие мужского генома.Другая статья Илменси и Хоппе имела еще больший резонанс.Авторы сообщили о пересадке ядер клеток внутренней клеточной массы бластоцисты в энуклеированные зиготы мышей и получении трех взрослых мышей (двух самок исамца), генетически идентичных донорской линии мышей. Введение ядер-доноров и удаление пронуклеусов из зиготы проводили за один прием, затем реконструированные яйцеклетки культивировали in vitro до стадии бластоцисты и пересаживали в матку самок. Из 16-ти пересаженных бластоцист три развились во взрослых животных. В следующей работе (1982) эти же авторы использовали в качестве доноров ядер клетки эмбрионов еще более поздних стадий (7 суток) и будто бы получили трех половозрелых мышей. Однако никто из работающих в том же направлении не смог добиться подобных результатов, и достоверность данных Илменси и Хоппе была вновь поставлена под сомнение. МакГрат и Солтер показали, что ядра 8-клеточных зародышей и клеток внутренней клеточной массы бластоцисты не обеспечивают развитие in vitro реконструированных яйцеклеток даже до стадии морулы, которая предшествует стадии бластоцисты. Небольшая часть (5%) ядер 4-клеточных зародышей дает возможность развиваться только до стадии морулы. В то же время 19% реконструированных яйцеклеток, содержащих ядра 2-клеточных зародышей, смогли достичь стадии морулы или бластоцисты.Эти и многие другие данные показывают, что в эмбриогенезе у мышей клеточные ядра рано теряют тотипотентность, что связано очевидно, с очень ранней активацией генома зародыша - уже на стадии 2-х клеток. У других млекопитающих, в частности, у кроликов, овец и крупного рогатого скота, активация первой группы генов в эмбриогенезе происходит позднее, на 8-16- клеточной стадии. Возможно поэтому первые значительные успехи в клонировании эмбрионов были достигнуты на других видах млекопитающих, а не на мышах. Тем не менее, работы с мышами, несмотря на их непростую судьбу, значительно расширили наши представления о методологии клонирования млекопитающих. Кролики, коровы и свиньи Американские исследонатели Стик и Робл, используя методику МакГрата и Солтера, получили 6 живых кроликов, пересадив ядра 8клеточных эмбрионов одной породы в лишенные ядра яйцеклетки кроликов другой породы. Фенотип родившихся полностью соответствовал фенотипу донора. Однако только 6 из 164 реконструированных яйцеклеток (3,7%) развились в нормальных животных. Это, конечно, очень низкий выход, практически не позволяющий рассчитывать на получение таким методом клона генетически идентичных животных. Ценность этой работы тем не менее в том. что она показала возможность клонирования эмбрионов кроликов.Работа с реконструированными яйцеклетками крупных домашних животных идет несколько по-другому. Их сначала культивируют не in vitro, a in vivo - в перевязанном яйцеводе овцы - промежуточного (первого) реципиента. Затем их оттуда вымывают и трансплантируют в матку окончательного (второго) реципиента - коровы или овцы соответственно, где их развитие происходит до рождения детеныша. Уиладсин предложил заключать реконструированные яйцеклетки в агаровый цилиндр, который он затем трансплантировал в перевязанный яйцевод овцы. По данным одних авторов реконструированные зародыши лучше развиваются в яйцеклетке, чем в культуральной среде, хотя некоторые исследователи получили неплохие результаты и при культивировании.Американцы Робл и его сотрудники, используя щадящий метод извлечения ядра без прокалывания мембраны яйцеклетки, предложенный МакГратом и Солтером, пересаживали в зиготы так называемые кариопласты - мужской и женский пронуклеусы вместе с окружающей их цитоплазмой, а также ядра 2-, 4- или 8- клеточных эбрионов коровы. Сначала зиготы центрифугировали чтобы освободить пронуклеусы от окружающих их гранул желтка, после чего ядра были хорошо видны под микроскопом, что значительно облегчало их удаление. При помощи манипулятора и заостренной стеклянной микропипетки извлекали один из бластомеров вместе с ядром из ранних зародышей и переносили его в энуклеированную зиготу. Реконструированные зародыши были заключены в агаровый цилиндр и пересажены в перевязанный яйцевод овцы. Через пять дней культивирования их вымывали, освобождали от агара и исследовали. Реконструктурированные зародыши в этой работе развивались только в тех случаях, когда в зиготы пересаживали пронуклеусы: 17% таких зародышей достигли стадии морулы или бластоцисты. Два зародыша были пересажены второму реципиенту - в матку коровы, и развитие их завершилось рождением живых телят. Если в качестве доноров использовали ядра 2-, 4- или 8-клеточных зародышей, то реконструированные яйцеклетки не развивались даже до стадии морулы.Позже были и более успешные работы. Уиладсин, в частности. сообщил, что ему удалось получить четырех генетически идентичных бычков холстейнской породы в результате пересадки в реципиентные яйцеклетки ядер бластомеров одного 32- клеточного зародыша. Автор утверждал, что большинство ядер сохраняет тотипотентность на 32-клеточной стадии, а значительная их часть даже на 64- клеточной стадии, обеспечивая нормальное развитие реконструированных яйцеклеток до стадии ранней бластоцисты в яйцеводе овцы. После пересадки в матку коров - окончательных реципиентов, как полагает автор, они могут и дальше нормально развиваться. Бондиоли и соавторы, используя в качестве доноров ядер 16-64-клеточные зародыши коров, трансплантировали 463 реконструированных зародыша в матку синхронизированных реципиентов, и было получено 92 живых теленка. Семь из них были генетически идентичны, представляя собой клон, полученный в результате пересадки ядер клеток одного донорского эмбриона.Таким образом, клеточные ядра зародышей крупного рогатого скота достаточно долго сохраняют тотипотентность и могут обеспечить полное развитие реконструированных яйцеклеток. Иначе говоря, методические трудности клонирования зародышей крупного рогатого скота практически решены. Но остается основная задача - найти донорские ядра, обладающие тотипотентностью, для клонирования взрослых животных.Клонированию эмбрионов свиней посвящена только одна небольшая работа. Скудность данных, видимо.и связана с определенными трудностями работы с этимобъектом.
|