
Клиническая анатомия наружного уха 2 страница
Иннервация слизистой оболочки барабанной полости. Происходит главным образом за счет барабанного нерва (п. tympani-cus) — ветвь языкоглоточного нерва (п. glossopharyngeus), анасто-мозирующего с веточками лицевого, тройничного нервов и симпатического внутреннего сонного сплетения. Сосцевидный отросток. Задний отдел среднего уха представлен сосцевидным отростком {processes mastoideus), в котором имеются многочисленные воздухоносные ячейки, соединенные с барабанной полостью через сосцевидную пещеру {antrum mas-toideum) и вход в пещеру {aditus ad antrum) в верхнезадней части надбарабанного пространства. У новорожденных сосцевидный отросток не развит и имеется в виде небольшого возвышения около барабанного кольца сверху и сзади, содержащего только одну полость — сосцевидную пещеру. Развитие сосцевидного отростка начинается на 2-м году жизни и завершается в основном к концу 6-го — началу 7-го года жизни. Сосцевидный отросток взрослого напоминает конус, опро- ■ '. чГ 333
Сосцевидная пещера — воздухоносная ячейка округлой формы, которая постоянно присутствует в сосцевидном отростке, независимо от формы и строения его. Является самым надежным анатомическим ориентиром практически при всех операциях на ухе. У грудных детей пещера расположена выше слухового прохода и довольно поверхностно (на глубине 2—4 мм), в дальнейшем постепенно смещается кзади и книзу. Крыша пещеры (tegmen antri) — костная пластинка, отделяющая ее от твердой мозговой оболочки средней черепной ямки. Строение сосцевидного отростка различается в зависимости от количества воздухоносных полостей в нем, их величины и расположения. Формирование этих полостей происходит путем замещения костномозговой ткани врастающим мукопериос-том. По мере роста кости количество сообщающихся с пещерой воздухоносных сосцевидных ячеек все время увеличивается. По характеру пневматизации различают пневматический, диплоический и склеротический типы строения сосцевидного отростка (рис. 5.8).
При пневматическом типе строения воздухоносные ячейки заполняют почти весь отросток и распространяются иногда даже на чешую височной кости, скуловой отросток, пирамиду. Обычно около пещеры образуется зона более мелких клеток, к периферии они становятся все более крупными, нередко при этом имеется большая верхушечная ячейка. Диплоический (спонгиозный, губчатый) тип строения характеризуется небольшим количеством воздухоносных ячеек, они располагаются преимущественно вокруг пещеры и представляют собой небольшие полости, ограниченные трабекулами. Склеротический (компактный) тип строения отростка является следствием нарушения обменных процессов или результатом перенесенных общих или местных воспалительных заболеваний. При этом сосцевидный отросток образован плотной костной тканью с отсутствием ячеек или их минимальным количеством. К задней поверхности сосцевидного отростка прилежит сигмовидный синус (sinus sigmoideus) — венозная пазуха, через которую осуществляется отток крови из мозга в систему яремной вены. Под дном барабанной полости сигмовидный синус образует расширение — верхнюю луковицу внутренней яремной вены. Синус представляет собой дупликатуру твердой мозговой оболочки и отграничен от клеточной системы сосцевидного отростка тонкой, но достаточно плотной костной пластинкой (lamina vitrea). Деструктивно-воспалительный процесс в сосцевидном отростке при заболеваниях среднего уха может привести к разрушению этой пластинки и проникновению инфекции в венозный синус. Предлежание синуса, когда он расположен близко к задней стенке слухового прохода, или латеропозиция (поверхностное расположение), представляет опасность его травмирования при операциях на ухе. На внутренней поверхности верхушки сосцевидного отростка имеется глубокая сосцевидная борозда (incisura mastoidea), где прикрепляется двубрюшная мышца. Через эту борозду гной иногда прорывается из сосцевидных ячеек под шейные мышцы. Кровоснабжение сосцевидной области. Осуществляется из системы наружной сонной артерии через заднюю ушную артерию (a. auricularis posterior), венозный отток — в одноименную вену, впадающую в наружную яремную вену (v. jugularis externa). Иннервация. Область сосцевидного отростка иннервируется чувствительными нервами из шейного сплетения: большим ушным нервом (п. auricularis magnus) и малым затылочным нервом (п. occipitalis minor). 5.1.3. Клиническая анатомия внутреннего уха Внутреннее ухо, или лабиринт, находится в толще пирамиды височной кости и состоит из костной капсулы и включенного в нее перепончатого образования, по форме повторяющего строение костного лабиринта (рис. 5.9).
Рис. 5.9. Костный лабиринт. 1 — окно преддверия; 2 — окно улитки; 3 — ампула латерального полукружного канала; 4 — передний полукружный канал; 5 — задний полукружный канал; 6 — улитка. • Костный лабиринт Различают три отдела костного лабиринта: • средний — преддверие (vestibulum); • передний — улитка {cochlea); • задний — система из трех полукружных каналов (canalis Латерально лабиринт является медиальной стенкой барабанной полости, в которую обращены окна преддверия и улитки, медиально граничит с задней черепной ямкой, с которой его соединяют внутренний слуховой проход (meatus acusticus internus), вестибулярный водопровод (aqueductus vestibuli), где проходят эндолимфатический проток {ductus endolymphaticus) к водопроводу улитки — aqueductus cochleae (каналец улитки — canaliculus cochleae). Улитка {cochlea). Представляет собой костный спиральный канал, имеющий у человека примерно два с половиной оборота вокруг костного стержня {modiolus), от которого внутрь канала отходит костная спиральная пластинка {lam. spiralis ossea). Улитка на разрезе имеет вид уплощенного конуса с шириной основания 9 мм и высотой 5 мм, длина спирального костного канала — около 32 мм. Костная спиральная пластинка вместе с перепончатой базилярной пластинкой, являющейся ее продолжением, и преддверной {рейсснеровой) мембраной {membrana vestibularis) образует внутри улитки самостоятельный канал — улитковый проток {ductus cochlearis), который делит канал улитки на два спиральных коридора — верхний и нижний. Верхний отдел канала — лестница преддверия {scala vestibuli), нижний — барабанная лестница {scala tympani). Лестницы изолированы друг от друга на всем протяжении, лишь в области верхушки улитки сообщаются между собой через отверстие — геликотре-ма {helicotrema). Лестница преддверия сообщается с преддверием, барабанная лестница граничит с барабанной полостью посредством окна улитки и не сообщается с преддверием. У основания костной спиральной пластинки имеется канал, в котором расположен спиральный узел улитки {gangl. spirale cochleae) — здесь находятся клетки первого биполярного нейрона слухового тракта. Костный лабиринт заполнен перилимфой, а находящийся в нем перепончатый лабиринт — эндолимфой. Преддверие. Это центральная часть лабиринта, филогенетически наиболее древняя. Преддверие представляет собой небольшую полость, внутри которой расположены два кармана: сферическое углубление {recessus sphericus) и эллиптическое углубление {recessus ellipticus). В первом, ближе к улитке, находится сферический мешочек {sacculus), во втором, примыкающем к полукружным каналам — эллиптический мешочек {utriculus). Передняя часть преддверия сообщается с улиткой через лестницу преддверия, задняя — с полукружными каналами. Полукружные каналы. Три полукружных канала расположены в трех взаимно перпендикулярных плоскостях: латеральный, или горизонтальный, {canalis semicircularis lateralis) находится под углом в 30° к горизонтальной плоскости; передний, или фронтальный вертикальный, канал {canalis semicircularis anterior) — во фронтальной плоскости; задний, или сагиттальный вертикальный, полукружный канал {canalis semicircularis posterior) располагается в сагиттальной плоскости. В каждом канале различают расширенную ампулу и гладкое колено, обращенные к эллиптическому углублению преддверия. Гладкие колена вертикальных каналов — переднего и заднего — слиты в одно общее колено. Таким образом, полукружные каналы соединены с эллиптическим углублением преддверия пятью отверстиями. Латеральная костная ампула подходит вплотную ко входу в пещеру, образуя его медиальную стенку. ♦ Перепончатый лабиринт Перепончатый лабиринт представляет собой замкнутую систему полостей и протоков, по форме в основном повторяющих костный лабиринт (рис. 5.10). Пространство между перепончатым и костным лабиринтом заполнено перилимфой. Это пространство очень незначительно в области полукружных протоков и несколько расширяется в преддверии и улитке. Перепончатый лабиринт подвешен внутри перилимфатического пространства при помощи соединительнотканных тяжей. Полости перепончатого лабиринта заполнены эндолимфой. Перилимфа и эндолимфа представляют гуморальную систему ушного лабиринта и функционально тесно связаны между собой. Перилимфа по своему ионному составу напоминает спинномозговую жидкость и плазму крови, эндолимфа — внутриклеточную жидкость. Биохимическое различие касается в первую очередь содержания ионов калия и натрия: в эндолимфе много калия и мало натрия, в перилимфе соотношение обратное. Перилим-фатическое пространство сообщается с подпаутинным посредством водопровода улитки (канальца улитки), эндолимфа нахо-
Рис. 5.10. Взаимоотношение костного и перепончатого лабиринтов. 1 — латеральный полукружный канал; 2 — передний и задний полукружные каналы; 3 — эллиптический мешочек; 4 — эндолимфатический мешок; 5 — сферический мешочек; 6 — улитка. дится в замкнутой системе перепончатого лабиринта и с жидкостями мозга сообщения не имеет. Считается, что эндолимфа продуцируется сосудистой полоской, а реабсорбция ее происходит в эндолимфатическом мешке. Избыточное продуцирование эндолимфы сосудистой полоской и нарушение ее всасывания могут привести к повышению внутрилабиринтного давления. С анатомической и функциональной точек зрения во внутреннем ухе выделяют два рецепторных аппарата: • слуховой, находящийся в улитковом протоке (ductus co- • вестибулярный, объединяющий мешочки преддверия (sac- Улитковый проток {ductus cochlearis). Располагается в улитке между лестницей преддверия и барабанной лестницей (рис. 5.11). На поперечном разрезе улитковый проток имеет треугольную форму: он образован преддверной (преддверная мембрана), барабанной (спиральная мембрана) и наружной стенками (рис. 5.12). Верхняя стенка обращена к лестнице преддверия и образована тонкой, состоящей из двух слоев плоских эпителиальных клеток преддверной (рейсснеровой) мембраной. Дно улиткового протока образует базилярная пластинка, отделяющая его от барабанной лестницы. Край костной спиральной пластинки посредством базилярной пластинки соединяется с противоположной стенкой костной улитки, где внутри улиткового протока располагается спиральная связ- ка __ Ug. spirale, верхняя часть которой, богатая кровеносными сосудами, называется сосудистой полоской (stria vascularis). Базилярная пластинка имеет обширную сеть капиллярных кровеносных сосудов и представляет образование, состоящее из поперечно расположенных эластичных волокон, длина и толщина которых увеличивается по направлению от основного завитка к верхушке. На базилярной пластинке, расположенной спиралевидно вдоль всего улиткового протока, лежит спиральный (кортиев) орган — периферический рецептор слухового анализатора (рис. 5.13). Спиральный орган состоит из нейроэпителиальных внутренних и наружных во-лосковых клеток, поддерживающих и питающих клеток (Дейтерса, Гензена, Клаудиуса), наружных и внутренних столбовых клеток, образующих кортиевы дуги. Кнутри от внутренних столбовых клеток расположен ряд внутренних волосковых клеток (их около 3500); снаружи от наружных столбовых клеток находится около 20 000 наружных волосковых клеток. Волосковые клетки синап-тически связаны с периферическими нервными волокнами, исходящими из биполярных клеток спирального узла улитки. Опорные клетки спирального
Рис. 5.11. Фронтальный разрез улитки. 1 — лестница преддверия; 2 — лестница улитки; 3 — улитковый проток; 4 — спиральный узел; 5 — улитковая часть пред-дверно-улиткового нерва.
Рис. 5.12. Расположение улиткового протока в базальном завитке улитки. 1 — лестница преддверия; 2 — барабанная лестница; 3 — улитковый проток.
Рис. 5.13. Спиральный (кортиев) орган. 1 — преддверная (рейсснерова) мембрана; 2 — волосковые клетки; 3 — спиральный узел; 4 — нервные волокна к волосковым клеткам; 5 — опорные клетки; 6 — покровная мембрана; 7 — сосудистая полоска. органа выполняют поддерживающую и трофическую функции. Между клетками спирального органа имеются внутриэпители-альные пространства, заполненные жидкостью, получившей название " кортилимфа". По химическому составу кортилимфа довольно близка эндолимфе, однако имеет и существенные отличия. Над волосковыми клетками спирального органа расположена покровная мембрана {membrana tectoria), которая так же, как и базилярная пластинка, отходит от края костной спиральной пластинки и нависает над базилярной пластинкой, поскольку наружный край ее свободен. Под ней также находится кортилимфа. Покровная мембрана состоит из протофибрилл, имеющих продольное и радиальное направление, в нее вплетаются волоски нейроэпителиальных наружных волосковых клеток. При колебаниях базилярной пластинки происходит натяжение и сжатие этих волосков, что приводит к преобразованию механической энергии колебаний стремени и жидкостей внутреннего уха в энергию электрического нервного импульса. В спиральном органе к каждой чувствительной волосковой клетке подходит только одно концевое нервное волокно, не дающее ответвлений к соседним клеткам, поэтому дегенерация нервного волокна приводит к гибели соответствующей клетки. Следует отметить, что существует афферентная и эфферентная иннервация чувствительных клеток спирального органа, осуществляющая центростремительный и центробежный поток. На внутренние волосковые клетки падает 95 % афферентной (центростремительной) иннервации. Наоборот, основной эфферентный поток направлен на наружные волосковые клетки. Перепончатые полукружные протоки {ductus semicirculares). 340 Находятся в костных каналах, повторяют их конфигурацию, но меньше их по диаметру, за исключением ампулярных отделов, которые почти полностью выполняют костные ампулы (рис. 5.14, а). Соединительнотканными тяжами, в которых проходят питающие сосуды, перепончатые протоки подвешены к эндос-ту костных стенок. Внутренняя поверхность протока выстлана эндотелием, в ампулах каждого из полукружных каналов располагаются ампулярные рецепторы, представляющие собой небольшой круговой выступ — ампулярный гребешок {crista ampul-laris), на котором размещены опорные и чувствительные ре-цепторные клетки, являющиеся периферическими рецепторами преддверной части преддверно-улиткового нерва. Среди рецепторных волосковых клеток выделяют более тонкие и короткие неподвижные волоски — стереоцилии, количество которых доходит до 50—100 на каждой чувствительной клетке, и один длинный и толстый подвижный волосок — киноцилий, располагающийся на периферии апикальной поверхности клетки. С волосковым аппаратом рецепторных клеток связывают процессы возбуждения вестибулярного аппарата. Движение эндолимфы при угловых ускорениях в сторону ампулы или гладкого колена полукружного протока приводит к раздражению нейроэпителиальных клеток. Предполагается, в частности, что изменение расстояния между киноцилиями и стерео-цилиями приводит к гипо- или гиперполяризации, следствием чего является увеличение или уменьшение потока импульсов от рецепторной клетки. В преддверии лабиринта имеются два перепончатых мешочка — эллиптический и сферический, в полости которых располагаются отолитовые рецепторы. Соответственно мешочкам они называются пятно эллиптического мешочка {macula utr-iculi) и пятно сферического мешочка {macula sacculi) и представляют собой небольшие возвышения на внутренней поверхности обоих мешочков, выстланных нейроэпителием (рис. 5.14, б). Этот рецепторный аппарат также состоит из опорных и чувствительных клеток. Волоски чувствительных клеток, переплетаясь своими концами, образуют сеть, которая погружена в желеобразную массу, содержащую большое число кристаллов карбоната кальция, имеющих форму октаэдров. Волоски чувствительных клеток вместе со статокониями (отолитами) и желеобразной массой образуют мембрану статоконий {отоли-товую мембрану). Среди волосков чувствительных клеток, так же как и в ампуллярных рецепторах, различают киноцилий и стереоцилии. Давление статокониев на волоски чувствительных клеток, а также смещение волосков при прямолинейных ускорениях является моментом трансформации механической энергии в электрическую в нейроэпителиальных волосковых клетках. Эллиптический и сферический мешочки соединены между собой посредством тонкого канальца — протока эллип-
Рис. 5.14. Схема вестибулярных рецепторов. а — ампулярный рецептор: 1 — просвет ампулы полукружного протока, 2 — ампулярный гребешок, 3 — просвет эллиптического мешочка, 4 — мембрана статоконий; б — статокониев рецептор: 1 — мембрана статоконий, 2 — рецеп-торные клетки. тического и сферического мешочков {ductus utriculosaccularis), который имеет ответвление — эндолимфатический проток (ductus endolymphaticus). Проходя в водопроводе преддверия, эндолимфатический проток выходит на заднюю поверхность пирамиды и там слепо заканчивается эндолимфатическим мешком (saccus endolymphaticus), представляющим собой расширение, образованное дупликатурой твердой мозговой оболочки.
Рис. 5.15. Кровоснабжение лабиринта. 1 — позвоночная артерия; 2 — бази-лярная артерия; 3 — передняя нижняя мозжечковая артерия; 4 — артерия лабиринта. Таким образом, вестибулярные сенсорные клетки расположены в пяти рецепторных областях: по одной в каждой ампуле трех полукружных протоков и по одной в двух мешочках преддверия каждого уха. В нервных рецепторах преддверия и полукружных протоков к каждой чувствительной клетке подходит не одно, а несколько нервных волокон, поэтому гибель одного из этих волокон не влечет за собой гибели клетки. Кровоснабжение внутреннего уха осуществляется через артерию лабиринта (a. labyrinthi), являющуюся ветвью базилярной артерии — a. basilaris (рис. 5.15). Во внутреннем слуховом проходе артерия лабиринта делится на три ветви: преддверную (г. vestibularis), преддвврно-улитковую (г. vestibulocochlearis) и улитковую (г. cochlearis). Особенности кровоснабжения лабиринта состоят в том, что ветви артерии лабиринта не имеют анастомозов с сосудистой системой среднего уха, преддверная мембрана лишена капилляров, а в области ампулярных и отолитовых рецепторов подэпителиальная капиллярная сеть находится в непосредственном контакте с клетками нейроэпителия; к нейро-эпителиальным волосковым клеткам спирального органа кровеносные сосуды не подходят, их питание осуществляется опосредованно через прилежащие к ним трофические клетки. Венозный отток из внутреннего уха идет по трем путям: венам водопровода улитки, венам водопровода преддверия и венам внутреннего слухового прохода. Иннервация внутреннего уха. Слуховой анализатор (рис. 5.16). Волосковые клетки спирального органа синапти-чески связаны с периферическими отростками биполярных клеток спирального узла улитки (gangl. spirale cochleae), расположенного в основании костной спиральной пластинки улитки. Центральные отростки биполярных нейронов спирального узла являются волокнами улитковой части преддверно-улиткового (VIII) нерва (п. vestibulocochlearis), который проходит через внутренний слуховой проход и в области мосто-мозжечкового треугольника входит в мост. На дне IV желудочка преддверно-
Рис. 5.16. Схема слухового анализатора. 1 — кора височной доли большого мозга; 2 — медиальное коленчатое тело; 3 — нижние холмики пластинки крыши среднего мозга (задние бугры четверохолмия); 4 — латеральная петля; 5 — улитковые ядра; б — верхние оливные ядра; 7 — спиральный узел; 8 — спиральный орган. улитковый нерв делится на два корешка: преддверный (верхний) и улитковый (нижний). Волокна улиткового корешка заканчиваются в латеральном углу ромбовидной ямки на клетках переднего улиткового ядра {nucl. cochlearis ventralis) и заднего улиткового ядра (nucl. co-chlearis dorsalis). Таким образом, клетки спирального узла вместе с периферическими отростками, идущими к нейроэпители-альным волосковым клеткам спирального органа, и централь- ными отростками, заканчивающимися в ядрах моста, составляют / нейрон слухового анализатора. На уровне улитковых ядер расположен ряд ядерных образований, принимающих участие в формировании дальнейших путей для проведения слуховых раздражений: ядра трапециевидного тела, верхнее оливное ядро, ядро латеральной петли. От переднего и заднего улитковых ядер начинается // нейрон слухового анализатора. Меньшая часть волокон этого нейрона идет по одноименной стороне, а большая часть перекрещивается и переходит на противоположную сторону моста, заканчиваясь в оливе и трапециевидном теле. Волокна III нейрона в составе латеральной петли идут к ядрам пластинки крыши среднего мозга (четверохолмия) и медиального коленчатого тела, откуда уже волокна IV нейрона после второго частичного перекреста направляются в височную долю большого мозга и оканчиваются в корковом отделе слухового анализатора, располагаясь преимущественно в поперечных височных извилинах (извилины Гешля). Проведение импульсов от кохлеарных рецепторов по обеим сторонам мозгового ствола объясняет то обстоятельство, что одностороннее нарушение слуха возникает только в случае поражения среднего и внутреннего уха, а также преддверно-улиткового нерва и его ядер в мосту. При одностороннем поражении латеральной петли, подкорковых и корковых слуховых центров импульсы от обоих кохлеарных рецепторов проводятся по непораженной стороне в одно из полушарий большого мозга и расстройства слуха может не быть. Слуховая система обеспечивает восприятие звуковых колебаний, проведение нервных импульсов к слуховым нервным центрам, анализ получаемой информации. Вестибулярный анализатор. Рецепторные клетки вестибулярного анализатора контактируют с окончаниями периферических отростков биполярных нейронов преддверного узла (gangl. vestibulare), расположенного во внутреннем слуховом проходе. Центральные отростки этих нейронов формируют преддверную часть преддверно-улиткового (VIII) нерва, который проходит во внутреннем слуховом проходе, выходит в заднюю черепную ямку и в области мостомозжечкового треугольника внедряется в вещество мозга. В вестибулярных ядрах продолговатого мозга, в дне IV желудочка, заканчивается / нейрон. Вестибулярный ядерный комплекс включает четыре ядра: латеральное, медиальное, верхнее и нижнее. От каждого ядра идет с преимущественным перекрестом // нейрон. Высокие адаптационные возможности вестибулярного анализатора обусловлены наличием множества ассоциативных путей ядерного вестибулярного комплекса (рис. 5.17). С позиций клинической анатомии важно отметить пять основных связей вестибулярных ядер с различными образованиями центральной и периферической нервной системы.
А Вестубулоспинальные связи. Начинаясь от латеральных ядер, в составе преддверно-спинномозгового пути они проходят в передних рогах спинного мозга, обеспечивая связь вестибулярных рецепторов с мышечной системой. А Вестибулоглазодвигателъные связи. Осуществляются через систему заднего продольного пучка: от медиального и нижнего ядер идет перекрещенный путь, а от верхнего ядра — неперекрещенный, к ядрам глазодвигательного нерва. А Вестибуловегетативные связи. Осуществляются от медиального ядра к ядрам блуждающего нерва, к ретикулярной формации, диэнцефальной области. А Вестибуломозжечковые пути. Проходят во внутреннем отделе нижней мозжечковой ножки и связывают вестибулярные ядра с ядрами мозжечка. А Вестибулокортикапьные связи. Обеспечиваются системой волокон, идущих от всех четырех ядер к таламусу. Прерываясь в последнем, далее эти волокна идут к височной доле большого мозга, где вестибулярный анализатор имеет рассеянное представительство. Кора большого мозга и мозжечок выполняют регулирующую функцию по отношению к вестибулярному анализатору. Посредством указанных связей реализуются разнообразные сенсорные, вегетативные и соматические вестибулярные реакции. 5.2. Клиническая физиология уха В ухе объединены два органа — слуха и равновесия, каждый из них содержит периферический рецепторный отдел анализатора. Рецепторы слуха и равновесия относятся к механорецепторам и характеризуются определенной общностью принципов функционирования. В то же время более молодой в филогенетическом и онтогенетическом отношениях слуховой аппарат уха отличается большей сложностью организации. Чувствительные элементы слухового анализатора в отличие от вестибулярных относятся к экстероцепторам. Рассмотрим особенности функционирования органа слуха и вестибулярного анализатора.
5.2.1. Функция органа слуха Слух человека является сложным процессом, для реализации которого необходимы проведение звуковой волны, преобразование ее в нервные импульсы, передача их в нервные Центры, анализ и интеграция звуковой информации. Соответственно различают такие функции органа слуха, как звукопро-ведение и звуковосприятие. Адекватным раздражителем органа
|
 кинутый вниз верхушкой. Верхней границей служит височная линия (linea temporalis), которая является продолжением скулового отростка и примерно соответствует уровню дна средней черепной ямки. Передней границей сосцевидного отростка является задняя стенка наружного слухового прохода, у задневерх-него края которого имеется выступ — spina suprameatum (ость Генле). Этот выступ соответствует проекции пещеры, расположенной на глубине примерно 2—2, 5 см от поверхности кости.
кинутый вниз верхушкой. Верхней границей служит височная линия (linea temporalis), которая является продолжением скулового отростка и примерно соответствует уровню дна средней черепной ямки. Передней границей сосцевидного отростка является задняя стенка наружного слухового прохода, у задневерх-него края которого имеется выступ — spina suprameatum (ость Генле). Этот выступ соответствует проекции пещеры, расположенной на глубине примерно 2—2, 5 см от поверхности кости.

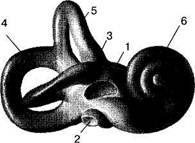



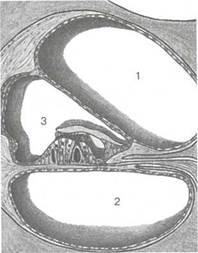



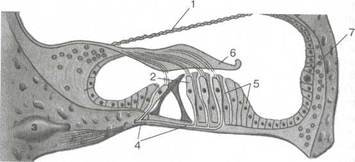






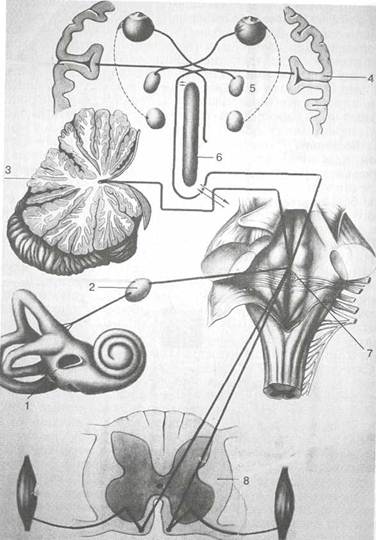
 Рис. 5.17. Схема ассоциативных связей вестибулярного анализатора. 1 — лабиринт; 2 — преддверный узел; 3 — мозжечок; 4 — кора полушарий большого мозга; 5 — ядра глазодвигательных нервов; б — ретикулярная формация; 7 — вестибулярные ядра в продолговатом мозге; 8 — спинной мозг.
Рис. 5.17. Схема ассоциативных связей вестибулярного анализатора. 1 — лабиринт; 2 — преддверный узел; 3 — мозжечок; 4 — кора полушарий большого мозга; 5 — ядра глазодвигательных нервов; б — ретикулярная формация; 7 — вестибулярные ядра в продолговатом мозге; 8 — спинной мозг.


