РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА
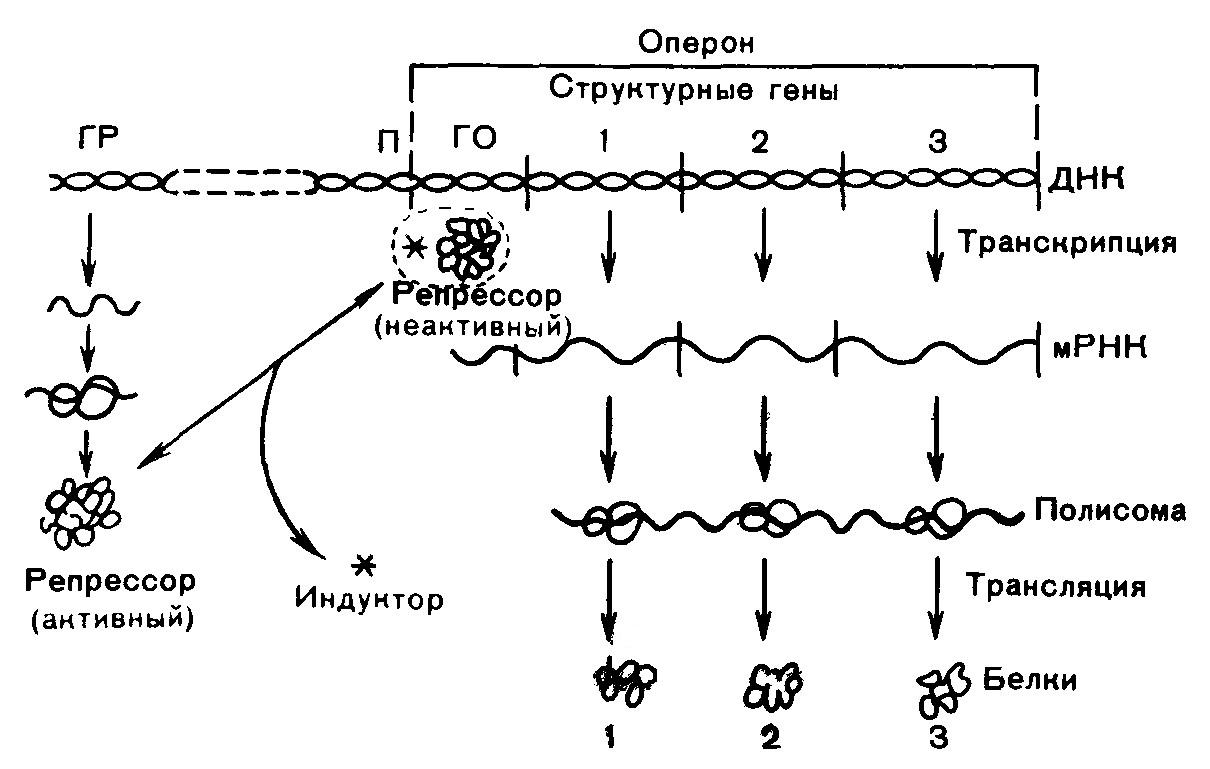
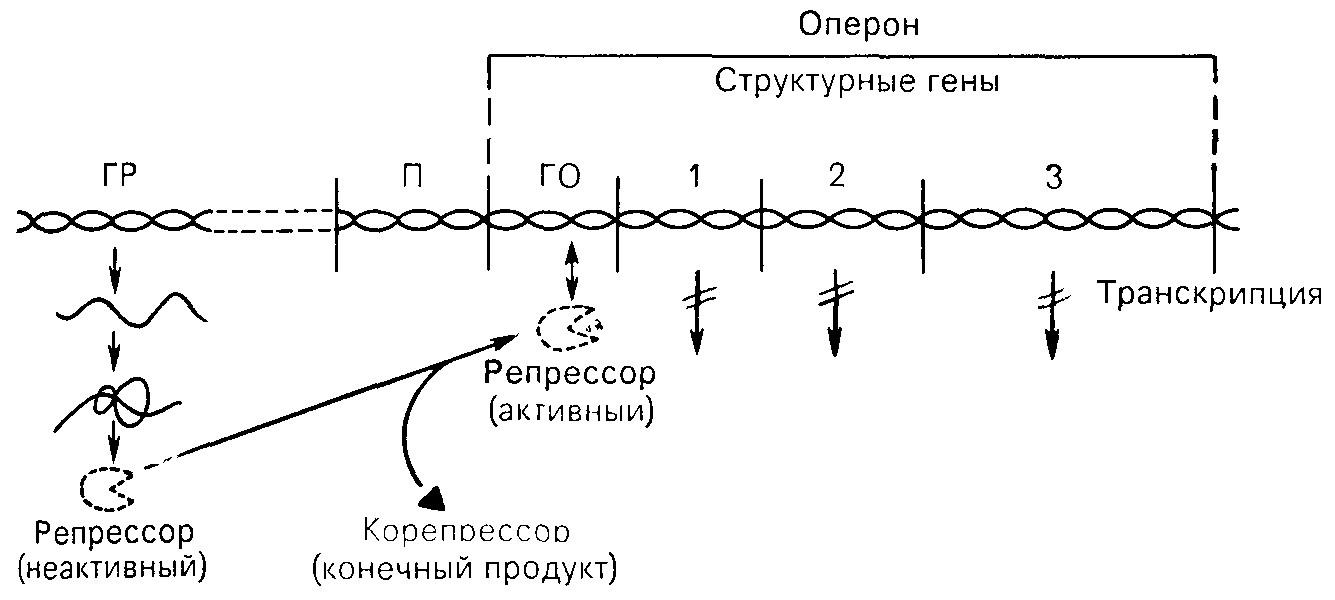
Концентрация многих белков в клетке непостоянна и изменяется в зависимости от состояния клетки и внешних условий. Это происходит в результате регуляции скоростей синтеза и распада белков. В клетках млекопитающих существуют два вида регуляции биосинтеза белков: - кратковременная, обеспечивающая адаптацию организма к изменениям окружающей среды; - длительная, стабильная, определяющая дифференцировку клеток и разный белковый состав органов и тканей. Регуляция на уровне транскрипции (образование первичного транскрипта) – наиболее распространенный механизм регуляции синтеза белков. Различают две формы регуляции – индукция синтеза (положительная регуляция) и репрессия синтеза (отрицательная регуляция). Понятия индукции и репрессии предполагают изменение скорости синтеза белка по отношению к исходному (базальному) уровню. Синтез в базальном состоянии называют конститутивным синтезом. Если скорость конститутивного синтеза белка высока, то белок регулируется по механизму репрессии синтеза, и, наоборот – при низкой базальной скорости бывает индукция синтеза. В генетическом аппарате клетки существуют опероны – отрезки ДНК, содержащие структурные гены определенных белков (цистроны), и регуляторные участки. Считывание генетического кода начинается с промотора, расположенного рядом с геном-оператором. Ген-оператор расположен на крайнем отрезке структурного гена. Он либо запрещает, либо разрешает репликацию мРНК на ДНК. Деятельность оперона контролирует ген-регулятор. Белок-репрессор осуществляет связь опероном и геном-регулятором. Репрессор образуется в рибосомах ядра на мРНК, синтезированной на гене-регуляторе. Он образует комплекс с геном-оператором и блокирует синтез мРНК, а, следовательно, и белка. Репрессор может связываться с низкомолекулярными веществами – индукторами, или эффекторами. После этого он теряет способность связываться с геном-оператором, ген-оператор выходит из-под контроля гена-регулятора, и начинается синтез мРНК. Это индукция синтеза (рис. 6). Наблюдается также эффект репрессии ферментов, когда концентрация ферментов в клетках снижается при увеличении содержания конечных продуктов цепи реакций синтеза (рис. 7). В этом случае репрессор синтезируется в неактивной форме и приобретают способность подавлять деятельность гена-оператора после образования комплекса с продуктом синтеза (корепрессором).
У эукариот преобладают положительные регуляторные механизмы. Основной регуляторной точкой является стадия инициации транскрипции. Регуляторные элементы, стимулирующие транскрипцию, называют энхансерами, а подавляющие ее – сайленсерами. Они могут избирательно соединяться с белками-регуляторами: энхансеры – с белками-индукторами, сайленсеры – с белками-репрессорами. Соединение регуляторных элементов с белками-регуляторами зависит от сигнальных молекул – гормонов, некоторых метаболитов и т.д. Знание особенностей структуры и функционирования рибосом прокариот и эукариот позволило разработать новые типы антибиотиков. Синтез нуклеиновых кислот и белков является ключевым процессом, необходимым для поддержания жизнедеятельности клетки. Если его каким-либо образом выключить - клетка погибнет. Существуют лекарственные препараты, нарушающие синтез пуриновых оснований и аминокислот, синтез нуклеиновых кислот, синтез белка на различных уровнях только в клетках бактерий.
|