Нейродегенерация и белки теплового шока
Суть клеточной реакции на тепловой шок состоит в том, что на стрессирующее воздействие клетка отвечает прекращением синтеза клеточных белков и начинает синтезировать строго ограниченный набор белковых молекул, которые были названы белками теплового шока (БТШ). Своим открытием белки теплового шока обязаны итальянскому ученому Ферруччо Ритосса. В 1962 г. помещая личинок Drosophila busckii в условия повышенной температуры, итальянский профессор Ферруччо Ритосса впервые обнаружил индукцию экспрессии БТШ, сопровождающуюся образованием 9 добавочных пуфов на политенных хромосомах (рис. 11) (De Maio, 2012). Политенными называют гигантские хромосомы, образованные стопками параллельно упакованных гомологичных нитей хроматина, которых может быть больше 1000. Пуф – вздутие на самой хромосоме, образующееся за счет разрыхления структуры хроматина в активном гене и накопления синтезирующихся молекул РНК (рис. 12). Впоследствии было показано, что пуффинг является видимым отражением активации транскрипции генов БТШ. Так же деформация политенных хромосом возникала в ответ на обработку клеток ферментными ядами, блокирующими окислительное фосфорилирование (Жимулев, 2003).
Рис. 11. Ферруччо Ритосса в своей лаборатории в начале 1960-х годов (по: De Maio, 2012).
Рис. 12. Динамика образования пуфа: а — нефункционирующая (спирализованная) хромосома, видна ее дискоидальная структура; б — начальное развитие пуфа, слабая деспирализация хромосомы, видны хромонемные нити; в — максимально развитый пуф, сильная деспирализация (по: Жимулев, 2003). Исследование БТШ происходило не только на дрозофиле. Пресноводная гидра Hydra oligactus неспособна синтезировать БТШ и погибает при незначительном прогревании. Близкий вид Hydra attenuata имеет нормальный синтез БТШ и выживает при таком же температурном воздействии (Bosch et al., 1988). Дальнейшие исследования показали, что синтез БТШ в ответ на тепловой стресс идентифицирован у широкого спектра организмов — от бактерии до человека (Schlesinger et al., 1982). Подтверждением защиты клетки БТШ от повреждений, вызванных гипертермией, является то, что организмы, не синтезирующие БТШ в ответ на тепловой шок, крайне термочувствительны. У ранних эмбрионов Drosophila melanogaster не происходит синтеза БТШ при действии ТШ (Foe, Alberts, 1985), поэтому период раннего эмбриогенеза D. melanogaster является удобной естественной моделью для изучения последствий действия ТШ в отсутствии синтеза БТШ. БТШ индуцируются не только на действие высоких температур, но и при разнообразных стрессорных воздействиях на организм. Например, помимо непосредственного участия всех БТШ в защите клеточных белков от разных видов денатурации и окисления, БТШ с помощью различных еще до конца не изученных механизмов участвуют в защите клеток от стресс-индуцируемого апоптоза, блокируя пути его активации и стабилизируя клеточные структуры. Подобный ответ на стрессорные воздействия был обнаружен у самых разных организмов, как эукариот, так и прокариот (Tanguay, 1988). В клетке синтезируется несколько фракций БТШ с различными молекулярными массами (Feder, Hofmann, 1999). Фракции БТШ разных организмов, приблизительно соответствуют друг другу по молекулярной массе. Общепринятой стала классификация, основанная на разделении БТШ по молекулярным массам на несколько семейств (Lindquist, 1986): 1) Семейство малых или низкомолекулярных БТШ (нмБТШ) с молекулярными массами от 10 до 30 кДа. Гены малых БТШ имеют бимодальную экспрессию. С одной стороны, они координированно активируются ТШ, с другой стороны, независимо экспрессируются на разных стадиях развития (Mason et al., 1984); 2) Группа высокомолекулярных БТШ с молекулярными массами от 40 и до 100 кДа. Ее основные семейства: 2.1. БТШ40 (40 кДа); 2.2. БТШ60 (58, 60 кДа); 2.3. БТШ70 (68, 70, 72, 73 кДа, кодируемые индуцибельными генами теплового шока); 2.4. БТШ90 (83, 87, 89 кДа); 2.5. БТШ100 (более 100 кДа – БТШ101, БТШ104, БТШ110). БТШ, усиливая или подавляя активность отдельных компонентов проведения внутриклеточного сигнала, обеспечивают либо сохранение целостности выполнения функций, либо приобретение новой функции этими компонентами, что позволяет осуществлять настройку компенсаторных механизмов на выполнение текущей потребности организма. Механизмы действия БТШ могут препятствовать нейродегенераци, которая в свою очередь является причиной развития НДЗ. НДЗ обладают рядом общих свойств. Эти заболевания характеризуются образованием аномальных клеточных и/или внеклеточных включений с последующей гибелью нейронов. Эта особенность позволила отнести НДЗ к группе конформационных болезней. В основе концепции «конформационных болезней мозга» лежит нарушение метаболизма и изменение конформации клеточных белков с их последующим накоплением и агрегацией в определенных группах нейронов (Takalo et al., 2013). Известны два белка, изменяющих структуру при НДЗ: α-синуклеин и тау-протеин (Holmes, Diamond, 2014). Белок α-синуклеин в норме присутствует в пресинаптических терминалях головного мозга. При НДЗ данный белок накапливается и формирует внутри глиальных клеток нитевидные структуры диаметром 20 — 40 нм (рис. 9).
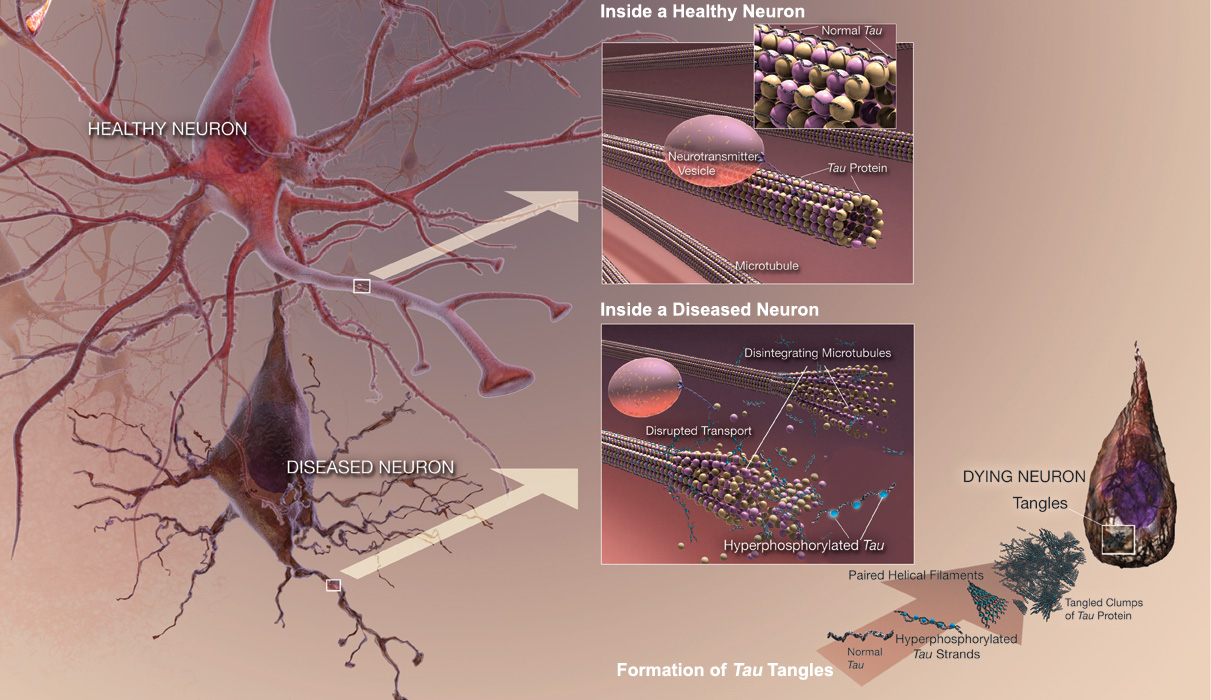
Рис. 9. Здоровые и больные нейроны при нарушении функций α-синуклеина (по: Lee et al., 2014). Тау-протеин представляет собой растворимый низкомолекулярный белок, играющий важную роль в процессе роста аксона и его функционировании. При НДЗ обнаруживаются его патологические формы, образующие нити, преобладающие в телах нейронов и аксонов (рис. 10).
Рис. 10. Здоровые и больные нейроны при нарушении функций тау-протеина (по: Bloom, 2014). Одно из современных объяснений возникновения этих отложений опирается на понимание того, как функционирует система универсального клеточного ответа на действие стрессорных факторов. При этом ответе происходит индукция белков стресса (более известных под названием белков теплового шока, БТШ), что может либо вызывать адаптацию организма (при кратковременном действии стрессорного фактора), либо приводить к гибели клеток (при длительном или слишком сильном его действии). БТШ выполняют шаперонную функцию в клетке – проверку правильности принятия третичной конформации белками (или фолдинга белков) при их синтезе и доставку в правильное место их функции в клетке. Известно, что при тепловом шоке и многих других видах стресса клеточные белки подвергаются денатурации. При этом гидрофобные области белковых молекул, в норме обращённые внутрь их структуры, экспонируются в водную среду цитоплазмы. Затем белки «слипаются» друг с другом гидрофобными участками, образуя нерастворимые агрегаты. Это приводит к нарушению функций большинства клеточных систем и гибели клетки. Было показано, что некоторые БТШ неспецифически взаимодействуют с гидрофобными участками денатурированных белков, способствуя восстановлению их нормальной вторичной и третичной структуры. В связи со способностью связываться с гидрофобными участками денатурированных и вновь синтезируемых белков, БТШ (и в первую очередь БТШ70) получили название «молекулярные шапероны» (от французского «chaperone» – компаньонка, дуэнья) (Lindquist, 1986). Молекулярные шапероны опосредуют активность различных киназ, рецепторов и факторов транскрипции (Rutherford, Zuker, 1994). Согласно амилоидной гипотезе, основной фактор, запускающий процесс нейродегенерации, - отложения β-амилоидного белка (Hardy, Allsop, 1991). β-амилоид является нормальным белком организма. Данный белок образуется в результате протеолитического процессинга из своего предшественника, названного предшественником бета-амилоида (ПБА), экспрессирующегося фактически во всех изученных к настоящему времени животных клетках. Современные исследования показали, что β-амилоид и его предшественники выполняют весьма важные физиологические функции. Однако, отложения β-амилоидного белка провоцируют апоптоз (Wang et al., 2012). Таким образом, белок может откладываться как вокруг нейронов с образованием бляшек, так и внутри нейронов с формированием нейрофибриллярных переплетений. Возможно, эти патологические процессы приводят к нарушениям синаптической передачи. Но не обнаружено четкой связи накопления бляшек с потерей нейронов (Schmitz et al., 2004). Отсутствие доказательств подобной связи приводит к тау-гипотезе, согласно которой каскад нарушений запускается гиперфосфорилированием тау-белка, приводящим к образованию нейрофибриллярных клубков, что нарушает организацию цитоскелета нейронов (Nakamura et al., 2013). Одним из ключевых моментов в формировании кофилин-актиновых палочек при ишемии мозга является АТФ-зависимое образование комплекса хронофинфосфатазы кофилина (cofilin phosphatase chronophin, CIN) и БТШ90. Активность CIN ингибируется образованием комплекса с БТШ90 (Huang et al., 2012). При АТФ-истощении CIN высвобождается из комплекса с БТШ90, что приводит к дефосфорилированию кофилина, его активации и иммобилизации кофилина и актина в составе палочек (Bernstein et al., 2006). Ингибитор БТШ90 – 17AAG, аналог галдономицина, также высвобождает CIN из комплекса с БТШ90, что вызывает активацию кофилина. Дефосфорилирование кофилина (активация кофилина) в нейронах происходит в течение ~20 мин после блокировки синтеза АТФ (Suurna et al., 2006). Формирование актин-кофилиновых включений, вызванное нарушением функционирования митохондрий, блокируется при действии цитохолазина D, который препятствует сборке актиновых филаментов и разбирает фибриллы актина до олигомеров, при этом количество глобулярного G-актина не изменяется (Goebel, Warlo, 1997). Таким образом, БТШ являются одним из механизмов предотвращения гибели клеток нервной системы из-за образования аномальных клеточных и/или внеклеточных включений. И, как следствие, БТШ препятствуют процессам нейродегенерации. Так же несомненна связь БТШ с компонентами актинового каскада при организации ответа клетки на тепловой шок, т.к. функционирование системы БТШ тесно связано с работой сигнального каскада ремоделирования актина и практически все клеточные белки хотя бы временно взаимодействуют с БТШ70 (например, в процессе синтеза белка на рибосоме).
|