Лекционный материал.
Хромосо́мы (др.-греч. χρῶμα — цвет и σῶμα — тело) — нуклеопротеидныеструктуры в ядре эукариотической клетки, в которых сосредоточена бо́льшая частьнаследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичнымпризнаком, для которого характерен относительно низкий уровень индивидуальной изменчивости. Хромосома образуется из единственной и чрезвычайно длинной молекулы ДНК, которая содержит линейную группу множества генов. Необходимыми функциональными элементами хромосомы эукариот являются центромера,теломеры и точки начала инициации репликации. Точки начала репликации (сайты инициации) и теломеры, находящиеся на концах хромосом, позволяют молекуле ДНК эффективно реплицироваться, тогда как в центромерах сестринские молекулы ДНК прикрепляются к митотическому веретену деления, что обеспечивает их точное расхождение по дочерним клеткам в митозе. Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят обактериальных или вирусных хромосомах. Поэтому, по мнению Д. Е. Корякова иИ. Ф. Жимулёва, более широким определением является определение хромосомы как структуры, которая содержит нуклеиновую кислоту и функция которой состоит в хранении, реализации и передаче наследственной информации. Хромосомы эукариот — это ДНК-содержащие структуры в ядре, митохондриях и пластидах. Хромосомы прокариот — это ДНК-содержащие структуры в клетке без ядра. Хромосомы вирусов — это молекула ДНК или РНК в составе капсида. Первые описания хромосом появились в статьях и книгах разных авторов в 70-х годах XIX века, и приоритет открытия хромосом отдают разным людям. Среди них такие имена, как И. Д. Чистяков (1873), А. Шнейдер (1873), Э. Страсбургер (1875),О. Бючли (1876) и другие. Чаще всего годом открытия хромосом называют 1882 год, а их первооткрывателем — немецкого анатома В. Флеминга, который в своей фундаментальной книге «Zellsubstanz, Kern und Zelltheilung» (нем.) собрал и упорядочил сведения о них, дополнив результатами собственных исследований. Термин «хромосома» был предложен немецким гистологом Г. Вальдейером в 1888 году. «Хромосома» в буквальном переводе означает «окрашенное тело», поскольку оснóвные красители хорошо связываются хромосомами. После переоткрытия в 1900 году законов Менделя потребовалось всего один-два года для того, чтобы стало ясно, что хромосомы при мейозе и оплодотворении ведут себя именно так, как это ожидалось от «частиц наследственности». В 1902 году Т. Бовери и в 1902—1903 годах У. Сеттон (Walter Sutton) независимо друг от друга первыми выдвинули гипотезу о генетической роли хромосом. Экспериментальное подтверждение этих идей и окончательное формулирование хромосомной теории было сделано в первой четверти XX века основателями классической генетики, работавшими в США с плодовой мушкой (D.melanogaster):Т. Морганом со своими учениками К. Бриджесом, А. Стёртевантом (A.H.Sturtevant) и Г. Мёллером. На основе своих данных они сформулировали «хромосомную теорию наследственности», согласно которой передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Эти выводы были опубликованы в 1915 году в книге «The mechanisms of mendelian heredity». В 1933 году за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине. Морфология метафазных хромосом
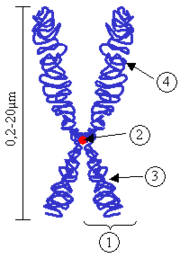
Схема строения хромосомы в метафазе митоза. 1 — хроматида; 2 —центромера; 3 — короткое плечо; 4 — длинное плечо. В ходе клеточного цикла облик хромосомы меняется. В интерфазе это очень нежные структуры, занимающие в ядре отдельныехромосомные территории, но не заметные как обособленные образования при визуальном наблюдении. В митозе хромосомы преобразуются в плотно упакованные элементы, способные сопротивляться внешним воздействиям, сохранять свою целостность и форму. Именно хромосомы на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа. Митотические хромосомы можно увидеть у любого организма, клетки которого способны делиться митозом, исключение составляют дрожжи S.cerevisiae, чьи хромосомы слишком малы. Обычно митотические хромосомы имеют размеры в несколько микрон. Например, самая большая хромосома человека хромосома 1 имеет длину около 7 — 8 мкм в метафазе и 10 мкм в профазе митоза. В зависимости от расположения центромеры различают три типа строения хромосом: акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах; субметацентрические хромосомы с плечами неравной длины; метацентрические хромосомы, у которых центромера расположена посередине или почти посередине. Эту классификацию хромосом на основе соотношения длин плеч предложил в 1912 году российский ботаник и цитолог С. Г. Навашин. Помимо вышеуказанных трёх типов С. Г. Навашин выделял ещё и телоцентрическиехромосомы, то есть хромосомы только с одним плечом. Однако по современным представлениям истинно телоцентрических хромосом не бывает. Второе плечо, пусть даже очень короткое и невидимое в обычный микроскоп, всегда присутствует. Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка, которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают различной длины и могут располагаться в различных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы, содержащие многократные повторы генов, кодирующих рибосомные РНК. У человека вторичные перетяжки, содержащие рибосомные гены, находятся в коротких плечах акроцентрических хромосом, они отделяют от основного тела хромосомы небольшие хромосомные сегменты, называемые спутниками. Хромосомы, обладающие спутником, принято называть SAT-хромосомами (лат. SAT (Sine Acid Thymonucleinico) — без ДНК). Уровни компактизации хромосомной ДНК Основу хромосомы составляет линейная макромолекула ДНК значительной длины. Например, в молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований. У эукариот существует высокоорганизованная система укладки молекул ДНК как в интерфазном ядре, так и в митотической хромосоме. Так, суммарная длина ДНК из одной клетки человека составляет величину порядка двух метров, в то время как типичноеядро человека, наблюдаемое только при помощи микроскопа, занимает объём около 110 мкм³, а митотическая хромосома человека в среднем не превышает 5 — 6 мкм. Надо отметить, что у эукариот в пролиферирующих клетках осуществляется постоянное закономерное изменение степени компактизации хромосом. Перед митозом хромосомная ДНК компактизуется в 105 раз по сравнению с линейной длиной ДНК, что необходимо для успешной сегрегации хромосом в дочерние клетки, в то время как в интерфазном ядре для успешного протекания процессов транскрипции и репликации хромосоме необходимо декомпактизоваться. При этом ДНК в ядре никогда не бывает полностью вытянутой и всегда в той или иной степени упакована. Так, расчётное уменьшение размера между хромосомой в интерфазе и хромосомой в митозе составляет всего примерно 2 раза у дрожжей и 4 — 50 раз у человека.
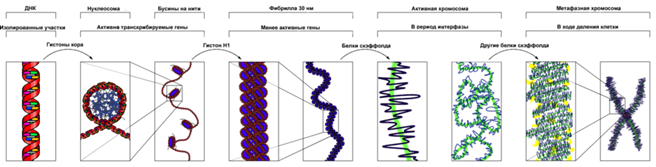
Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК, необходимое для размещения её в ядре. При этом надо оставить доступными определённые последовательности ДНК для регуляторных факторов и ферментов транскрипции. Эти задачи решаются на уровне упаковки ДНК в хроматин, которая происходит в несколько этапов. Наиболее изученными является три первых уровня упаковки: (1) накручивание ДНК на нуклеосомы с образованием нуклеосомной нити диаметром 10 нм, (2) компактизация нуклеосомной нити с образованием так называемой 30-нм фибриллы и (3) сворачивание последней в гигантские (50 — 200 тысяч п. н.) петли, закреплённые на белковой скелетной структуре ядра — ядерном матриксе. Одним из самых последних уровней упаковки в митотическую хромосому некоторые исследователи считают уровень так называемой хромонемы, толщина которой составляет около 0,1 — 0,3 мкм[21]. В результате дальнейшей компактизации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз). Анеуплоидия При анеуплоидии происходит изменение числа хромосом в кариотипе, при котором общее число хромосом не кратно гаплоидному хромосомному набору n. В случае утраты одной хромосомы из пары гомологичных хромосоммутантов называют моносомиками, в случае одной дополнительной хромосомы мутантов с тремя гомологичными хромосомами называют трисомиками, в случае утраты одной пары гомологов — нуллисомиками]. Анеуплоидия по аутосомным хромосомам всегда вызывает значительные нарушения развития, являясь основной причинойспонтанных абортов у человека. Одной из самых известных анеуплоидий у человека является трисомия по хромосоме 21, которая приводит к развитию синдрома Дауна[1]. Анеуплоидия характерна для опухолевых клеток, особенно для клеток сóлидных опухолей. Полиплоидия Изменение числа хромосом, кратное гаплоидному набору хромосом (n), называется полиплоидией. Полиплоидия широко и неравномерно распространена в природе. Известны полиплоидные эукариотические микроорганизмы —грибы и водоросли, часто встречаются полиплоиды среди цветковых, но не среди голосемянных растений. Полиплоидия клеток всего организма у многоклеточных животных редка, хотя у них часто встречаетсяэндополиплоидия некоторых дифференцированных тканей, например, печени у млекопитающих, а также тканей кишечника, слюнных желёз, мальпигиевых сосудов ряда насекомых. Хромосомные перестройки Хромосомные перестройки (хромосомные аберрации) — это мутации, нарушающие структуру хромосом. Они могут возникнуть в соматических и зародышевых клетках спонтанно или в результате внешних воздействий (ионизирующее излучение, химические мутагены, вирусная инфекция и др.). В результате хромосомной перестройки может быть утрачен или, наоборот, удвоен фрагмент хромосомы (делеция и дупликация, соответственно); участок хромосомы может быть перенесён на другую хромосому (транслокация) или он может изменить свою ориентацию в составе хромосомы на 180° (инверсия). Существуют и другие хромосомные перестройки. В каждой ядросодержащей соматической клетке человека содержится 23 пары линейных хромосом, а также многочисленные копии митохондриальной ДНК. В нижеприведённой таблице показано число генов и оснований в хромосомах человека. Вопросы:
ТОО «Есикский медицинский колледж» г.Есик
|