Проміжний обмін нуклеїнових кислот
Біосинтез нуклеїнових кислот
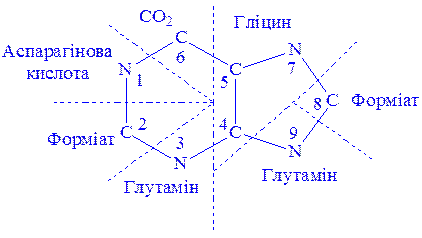
Матеріалом для синтезу нуклеїнових кислот у клітинах можуть бути екзогенні продукти гідролітичного розщеплення ДНК і РНК їжі. Частину матеріалів для цих процесів клітини одержують у вигляді продуктів вуглеводного, ліпідного і білкового обміну. З трьох складових частин нуклеотидів – пуринових і піримідинових основ, пентоз і фосфатів – дві останніх завжди є в клітині як продукти проміжного обміну. Пуринові і піримідинові основи клітина одержує в результаті всмоктування поживних речовин або біосинтезу з більш простих сполук. Біосинтез пуринових нуклеотидів. Використання мічених атомів (15N і 13C) показало, що в утворенні пуринового ядра бере участь ряд речовин. Так, два атоми азоту пуринового ядра походять від амідної групи глутаміна, по одному атому азоту – від гліцина і аспарагінової кислоти, атоми вуглецю – від форміата, гліцина і вуглекислого газу (рис.2).
Рис. 2. Походження атомів пуринового ядра
Біосинтез пуринових нуклеотидів відбувається поетапно. 1. Рибозо-5-фосфат під впливом ферменту рибозофосфат-пірофосфокінази взаємодіє з АТФ, що призводить до утворення a-5-фосфорибозил-1-пірофосфата (ФРПФ):
2. ФРПФ під впливом ферменту амідофосфорибозил-трансферази амінується за рахунок глутаміна, утворюючи b-5-фосфорибозил-1-амін:
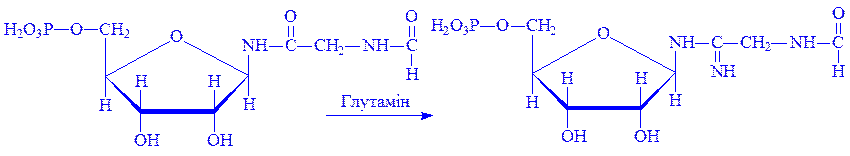
3. b-5-фосфорибозил-1-амін під впливом ферменту фосфорибозилгліцинамід-синтетази вступає в реакцію з гліцином, утворюючи гліцинамідрибонуклеотид:
4. Гліцинамідрибонуклеотид за участю ферменту фосфорибозилгліцинамід-форміл-трансферази вступає в реакцію з N5, Н10-метилентетрагідрофолієвою кислотою, утворюючи N-форміл-гліцинамід-рибонуклеотид:
5. Далі під впливом ферменту фосфорибозилформіл-гліцинамідинсинтетази, речовина, що утворилася вступає в реакцію з глутаміном і перетворюється на формілгліцинамідин-рибонуклеопротеїд (форміл-ГАМ):
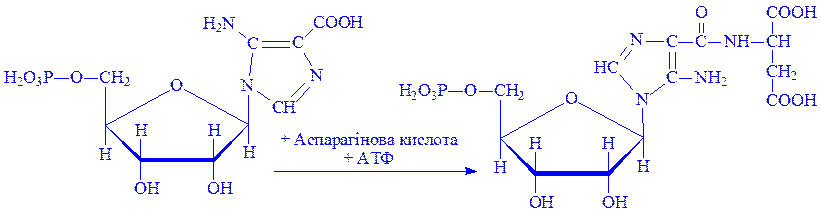
6. Форміл-ГАМ взаємодіє з АТФ під впливом ферменту фосфорибозиламіноімідазол-синтетази у присутності іонів Mg2+. Відбувається замикання кільця, що призводить до утворення 5-аміноімідазолрибонуклеотида:
7. Останній під впливом фосфорибозиламіноімідазол-карбоксилази карбоксилюється і перетворюється на рибонуклеотид 5-аміноімідазол-4-карбонової кислоти:
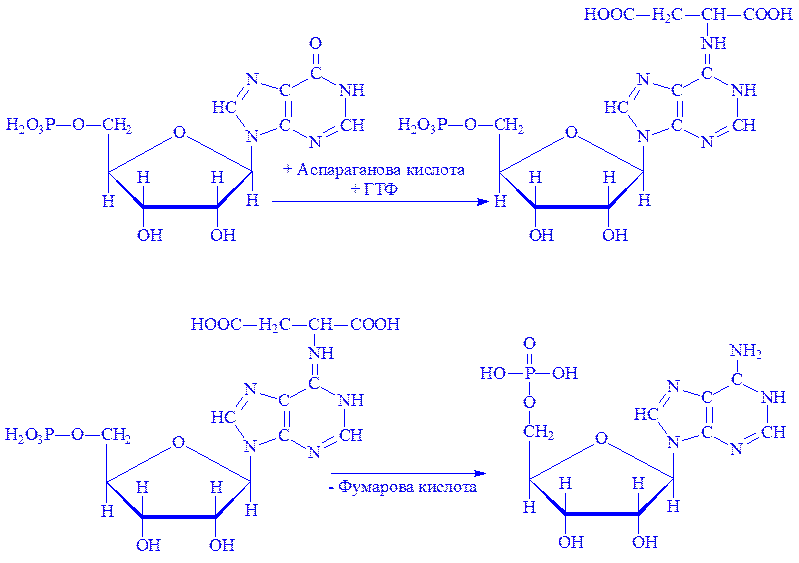
8. Сполука, яка утворилася, за участю ферменту 5-аміноімідазолсукцинокарбоксамід-синтетази і АТФ, взаємодіє з аспарагіновою кислотою:
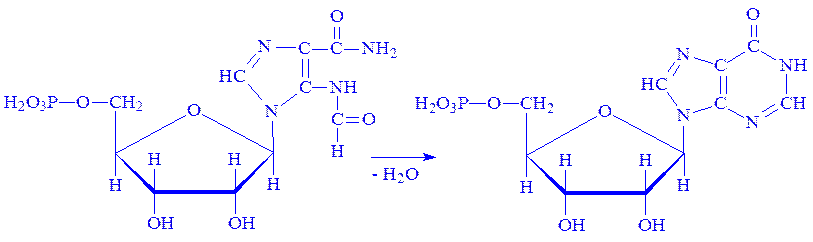
9. 5-Аміноімідазол-4-N-сукцинокарбоксамід-рибонуклеотид реагує під впливом ферменту фосфорибозилгліцинамід-формілтрансферази з N10-формілтетрагідрофолієвою кислотою:
10. 5-Формамідімідазол-4-карбоксамідрибонуклеотид під впливом ферменту ІМФ-циклогідролази втрачає молекулу води, циклізується і перетворюється на інозинову кислоту – попередник аденілової і гуанілової кислот:
Далі інозинова кислота під впливом ферменту 5-аміноімідазол-сукцинокарбоксамід-синтетази взаємодіє з аспарагіновою кислотою з утворенням аденілоянтарної кислоти, молекула якої надалі розпадається на аденілову і фумарову кислоти. Донором енергії є ГТФ:
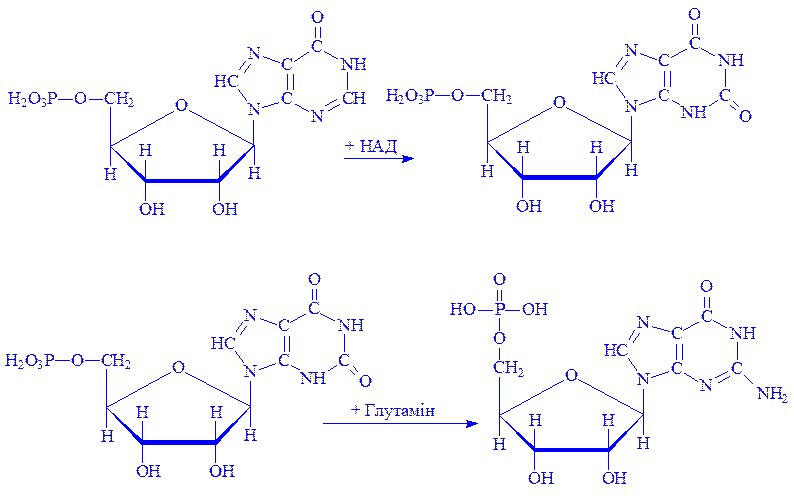
При утворенні молекули гуанілової кислоти інозинова кислота окислюється до ксантозин-5'-фосфата, який після взаємодії з глутаміном перетворюється на гуанілову кислоту:
Для біосинтезу полінуклеотидів потрібні макроергічні зв'язки. Вони утворюються за рахунок АТФ або іншого донатора енергії. Реакція протікає в дві стадії під впливом ферментів нуклеозидмонофосфат- і нуклеозиддифосфаткіназ:
ГМФ + АТФ ® ГДФ + АДФ; ГДФ + АТФ ® ГТФ + АДФ. Біосинтез піримідинових нуклеотидів. Початковою речовиною для синтезу піримідинових нуклеотидів є уридин-5'-монофосфат, який синтезується з урацила, пентози і фосфату. Пентози і фосфат завжди є в клітинах і міжклітинній рідині. Піримідинове кільце виникає з амінокислот, вуглекислого газу і форміата. Використання мічених атомів показало, що головним постачальником азоту є аспарагінова кислота і глутамін, вуглецю – CO2 і форміат. Процес утворення нуклеотидів протікає поетапно. 1. Спочатку під впливом ферменту карбамоїлфосфатсинтази за наявності NH3 і CO2 енергії АТФ утворюється карбамоїл фосфат:
2. Під впливом ферменту аспартат-карбамоїлтрансферази карбамоїл фосфат взаємодіє з аспарагіновою кислотою, що призводить до утворення карбамоїласпартата:
3. У результаті взаємодії аміногрупи карбамоїласпартата з карбоксильною групою виникає циклічна сполука – дигідрооротова кислота, яка під впливом ферменту дегідрогенази перетворюється на оротову кислоту:
4. Під впливом оротидин-5'-фосфат-пірофосфорилази оротова кислота вступає в реакцію з 5-фосфорибозил-1-пірофосфатом, утворюючи оротидин-5-фосфат, який декарбоксилується і перетворюється на уридин-5'-фосфат:
5. Уридин-5'-фосфат – попередник інших піримідинових нуклеотидів. Для цього УМФ під впливом кіназ за рахунок АТФ перетворюється в УДФ і УТФ:
УМФ + АТФ ® УДФ + АДФ; УДФ + АТФ ® УТФ + АДФ.
6. Перетворення урацила в цитозин відбувається на рівні нуклеозидтрифосфатів під впливом ЦТФ-синтетази:
УТФ + NH3 + АТФ ® ЦТФ + АДФ + Фн де Фн – фосфат неорганічний.
7. Перетворення урацила в тимін відбувається внаслідок реакції метилювання з S-аденозилметіоніном (CH3–R) утворюється ТТФ:
УТФ + CH3–R + АТФ ® ТТФ + АДФ + Фн + R
Перетворення піримідинових і пуринових рибонуклеотидів у дезоксирибонуклеотиди відбувається в результаті реакції відновлення, наприклад:
ТТФ Матричний синтез нуклеїнових кислот. Біосинтез ДНК відбувається під час інтерфази мітотичного поділу клітини. ДНК концентрується в хромосомах і хроматині. Синтез ДНК. Оскільки у молекулах ДНК закодована вся генетична інформація організму, синтез і відтворення їх є необхідним і життєво-важливим процесом. Процес синтезу ДНК називається реплікацією (копіювання, самоподвоєння), оскільки передача інформації відбувається в межах одного виду нуклеїнових кислот (від ДНК до ДНК). Реплікація відбувається у процесі росту і нормального поділу ДНК-вмісних структур (ядер, мітохондрій, пластид в еукаріот), а також у процесі поділу бактеріальних клітин та під час розмноження вірусів. Гіпотезу щодо механізму реплікації запропонували Д. Уотсон і Ф. Крік після того, як було встановлено, що молекула ДНК складається з двох взаємокомплементарних полінуклеотидних ланцюгів. Суть даного механізму полягає в тому, що синтез ДНК здійснюється на одноланцюговій матриці, роль якої виконує ланцюг молекули ДНК, що реплікується. Подвійна спіраль ДНК при цьому розкручується і на кожному з ланцюгів добудовується новий комплементарний ланцюг (утворюється репліка). Пізніше було встановлено, що для ДНК характерним є напівконсервативний механізм реплікації (від англ. conserve – зберігати) – у кожній новій синтезованій молекулі один ланцюг „старий”, одержаний з вихідної материнської ДНК, а другий – „новий”, синтезований (дочірній), тобто реплікація проходить так, що два ланцюги – вихідний і реплікований, сполучаючись між собою, утворюють наступне покоління молекул ДНК. Внаслідок цього процесу з однієї вихідної молекули ДНК утворюються дві молекули – точні копії, репліки, абсолютно ідентичні одна одній і вихідній матричній молекулі ДНК (рис. 3).
Рис. 10. Схема реплікації ДНК
Як якісний, так і кількісний вміст залишків нуклеотидів у полінуклеотидних ланцюгах материнської і дочірніх ДНК повністю співпадає. Напівконсервативний механізм синтезу ДНК відіграє важливу роль у забезпеченні видоспецифічної передачі генетичної інформації від покоління до покоління і є запорукою збереження сталості видів організмів. Згідно з дослідженнями А. Корнберга, вихідними сполуками для синтезу ДНК є вільні дезоксирибонуклеозид-5-трифосфати азотистих основ (аденіну, гуаніну, цитозину, тиміну), які під впливом відповідних ферментів сполучаються між собою з утворенням полінуклеотидних ланцюгів. Загальну схему синтезу ДНК можна подати так:
Реплікація ДНК – багатоступінчастий процес, для здійснення якого необхідні певні умови і наявність чотирьох видів рибо- і дезоксирибонуклеозид-5-трифосфатів, матриці у вигляді дволанцюгової ДНК, ферментів і білкових речовин, затравки (праймера), іонів (Mg2+, Mn2+). Матрицею служить молекула ДНК, яка реплікується. Разом з дим для здійснення реплікації необхідно близько 20 реплікативних факторів, що називаються ДНК-репліказною системою, або репліосомою. Важливими компонентами ДНК-репліказної системи є ферменти синтезу ДНК. У процесі реплікації беруть участь ферменти ДНК-залежні ДНК-полімерази (І, II, ІІІ) РНК-полімерази, ДНК-ліази, екзонуклеази, ДНК-лігази. Кожен з цих ферментів забезпечує здійснення певних етапів синтезу ДНК і часто виявляє кілька видів ферментативної активності. Зокрема, вважають, що ДНК-полімераза III, яка відіграє головну роль у процесі реплікації, має 5' ® 3' полімеразну і 3' ® 5' екзонуклеазну активності, ДНК-полімераза І має три види ферментативної активності: 5¢ ® 3' полімеразну, 3' ® 5' екзонуклеазну і 5' ® 3' екзонуклеазну. Цей фермент забезпечує розщеплення PHK затравки на матриці ДНК та процеси репарації пошкоджених ділянок молекули. ДНК-полімераза II за властивостями подібна до ДНК-полімерази І, однак для неї не характерна 5' ® 3' екзонуклеазна активність. Функції цього ферменту в клітині вивчені не достатньо. РНК-полімераза забезпечує синтез затравного ланцюга PHK (праймера) на ДНК-матриці. Важливими компонентами ДНК-репліказної системи є також білкові речовини, які забезпечують протікання ряду необхідних етапів синтезу ДНК, зокрема таких, як: 1) „впізнавання” місця початку реплікації ДНК ферментами ДНК-полімеразами; 2) локальне розкручування дуплексу ДНК і вивільнення одноланцюгових матриць; 3) стабілізація утвореної структури; 4) синтез затравних ланцюгів (праймерів) для ініціації ДНК-полімерази III; 5) переміщення реплікативної вилки в ході копіювання; 6) закінчення синтезу та утворення нативної конформації молекули ДНК. Ці процеси забезпечуються ДНК-зв’язуючими білками (ДЗБ), ДНК-розкручуючими білками (хеліказами), ДНК-стабілізуючими білками (гіразами) тощо. Слід зазначити, що реплікація ДНК має певні особливості, зокрема, напівконсервативний механізм, одночасність синтезу обох ланцюгів, двонапрямленість реплікації, синтез дочірніх ланцюгів у напрямку 5' ® 3¢, наявність праймера з вільною 3' –ОН групою дезоксирибози, фрагментарний характер реплікації. Процес реплікації ДНК включає три взаємозалежні і взаємозв'язані етапи: ініціацію, елонгацію і термінацію. Ініціація. Цей етап синтезу ДНК об'єднує процеси, які забезпечують початок синтезу полінуклеотидного ланцюга. Суть їх полягає у формуванні реплікативної вилки та синтезі праймера. Оскільки для забезпечення напівконсервативного механізму синтезу ДНК необхідна наявність одноланцюгової матриці, важливою умовою початку реплікації є дестабілізація трьохмірної структури ДНК та розкручування її подвійної спіралі з утворенням реплікативної вилки (рис. 4).
Рис. 4. Реплікативна вилка
В еукаріот, на відміну від прокаріот, утворюється багато стартових точок реплікації – від 10 до 100 тисяч. Кожна з них розміщується в окремих ділянках полінуклеотидного ланцюга, і реплікація їх проходить незалежно. Такі самостійні одиниці реплікації називаються репліконами. У бактерій роль реплікону виконує хромосома. Отже, в еукаріот молекула ДНК – це полірепліконна система, що містить велику кількість репліконів, з середини яких у двох напрямках рухаються реплікативні вилки. ДНК при утворенні реплікативної вилки розкручується не по всій довжині, а невеликими ділянками – відрізками, що містять до 2 тисяч неспарепих основ. Утворення реплікативної вилки забезпечується білковими речовинами, які визначають точку старту реплікації та зумовлюють розкручування подвійної спіралі ДНК. У клітинах еукаріот ці процеси забезпечуються комплексною дією ДНК-розкручуючих білків. Так, топоізомерази руйнують суперспіралізовану структуру ДНК хроматину, свівелази або релаксуючі білки, зумовлюють розрив одного з ланцюгів ДНК, внаслідок чого стає можливим розкручування двоспіральної структури ДНК за участю хеліказ. У результаті поетапної дії вищезазначених факторів двоспіральна молекула ДНК розкручується й утворюється дві одноланцюгові матриці. Оскільки в двоспіральній молекулі ДНК ланцюги антипаралельні – один з них має напрямок 5¢ ® 3', а інший – 3' ® 5', то після розкручування дуплексу утворюються одноланцюгові антипаралельні матриці. В зв'язку з тим, що реплікація за участю ДНК-полімерази III може проходити лише в напрямку 5' ® 3¢ напрямок реплікації одного ланцюга співпадає з напрямком реплікативної вилки, цей ланцюг називається ведучим, а ланцюг, реплікація якого протилежна напрямку реплікативної вилки, – тим, що запізнюється, другорядним. Після розкручування подвійної спіралі та утворення реплікативної вилки наступає друга стадія ініціації – синтез праймера, або затравного ланцюга, PHK на структурі ДНК-матриці. Необхідність цього процесу зумовлена тим, що ДНК-полімераза III не здатна самостійно розпочати синтез ДНК на структурі матриці з використанням вільних нуклеозидтрифосфатів. Тому для ініціації синтезу ДНК необхідна затравка, яка б містила вільну 3'–ОН групу, за якою може здійснюватись ДНК-полімеразна ІІI реакція. Встановлено, що функцію затравки виконують невеликі олігонуклеотидні фрагменти PHK, комплементарні ДНК-матриці. Затравка в середньому містить 8 – 10 нуклеотидів. Синтез затравного полінуклеотидного ланцюга каталізує мультиферментний комплекс, так звана „праймосома”. До складу праймосоми входять ферменти – специфічна ДНК-залежна РНК-полімераза (ДНК-праймаза), ДНК-залежна АТФ-аза, а також білкові фактори: ДНК-зв'язуючий білок (білок В), білок n¢ тощо. ДНК-зв'язуючий білок визначає місце старту, з якого праймаза розпочинатиме синтез затравного ланцюга в напрямку 5' ® 3'. Праймосома рухається по ланцюгах матриці і синтезує праймер. Цей процес розпочинається із приєднання до стартової точки ведучого полінуклеотидного ланцюга матриці ДНК-зв¢язуючого білка. Далі за участю праймази проходить синтез затравкого ланцюга PHK, комплементарного матриці ДНК. Після закінчення синтезу на ведучому ланцюгу праймосома переміщується на ланцюг, що запізнюється, й ініціює синтез праймера на ньому. По ланцюгу ДНК, що запізнюється, праймосома рухається до наступного місця ініціації в напрямку, протилежному напрямку синтезу ДНК, але однонапрямленому з реплікативною вилкою, синтезує затравку і знову переміщується вперед. Враховуючи це, білок В називають мобільним промотором реплікації. Енергія для руху праймосоми забезпечується завдяки активності АТФ-аз білка n'. Отже, в результаті дії праймосоми синтезується РНК-затравка на структурі ДНК-матриці. На 3'–ОН кінці затравки замість праймосоми приєднується ДНК-залежна ДНК-полімераза III, яка використовує 3¢–ОН групу для продовження копіювання матричної молекули ДНК. Синтезом праймерів завершується процес ініціації. Елонгація, або подовження полінуклеотидного ланцюга, включає два процеси: реплікацію ділянок материнської ДНК і сполучення синтезованих ділянок між собою. Ці процеси забезпечуються ДНК-полімеразою III, яка виявляє полімеразну та нуклеотидилтрансферазну активність. Отже, в цьому випадку ДНК-полімераза III діє на два субстрати: на спарений з матрицею кінець затравки, яка містить вільну 3'–ОН групу рибози, і на дезоксирибонуклеозид-5-трифосфати (дРТФ). Хімізм реакції поліконденсації полягає в нуклеофільній атаці a-атома фосфату дРТФ 3'–ОН групою, яка міститься на кінці затравки. В результаті реакції дезоксирибокуклеозид-5-монофосфат приєднується за місцем атома водню гідроксильної групи з утворенням 3' ® 5'-фосфодіефірного зв'язку і вивільненням пірофосфату. Продукт реакції містить вільну 3'–ОН групу, за якою знову повторюється аналогічна реакція, що дістала назву „ріст з хвоста”. В результаті багаторазового повторення цієї реакції здійснюється поступовий ріст нового полінуклеотидного ланцюга, комплементарного матричному ланцюгу ДНК. Дослідженнями, японський вчений Оказакі в 1968 р., встановив, що синтез полінуклеотидів ДНК на матричній ДНК проходить не безперервно, а окремими фрагментами, які дістали назву фрагментів Оказакі. Утворюються вони внаслідок приєднання до 3'–ОН групи праймерів 1000 – 2000 дезоксирибонуклеотидних залишків. Послідовність нуклеотидів синтезованих фрагментів комплементарна нуклеотидній послідовності відповідної ділянки ланцюга матриці. Поступове нарощування фрагментів Оказакі триває доти, поки не досягне місця, де розташована нова затравка, з якої розпочинається ріст нового фрагменту полінуклеотидного ланцюга. Тепер вважають, що фрагментарний механізм реплікації властивий в основному полінуклеотидному ланцюгу, що запізнюється. Вважають, що для ведучого ланцюга фрагментація не характерна. За даними Оказакі, в живих клітинах має місце човниковий механізм реплікації. Суть його полягає в тому, що спочатку реплікація відбувається в напрямку 5' ® 3' під час просування вперед по одному ланцюгу, а потім у напрямку 5' ® 3' продовжується в протилежному напрямку по другому полінуклеотидному ланцюгу матричної ДНК. Інакше кажучи, один полінуклеотидний ланцюг синтезується в напрямку реплікативної вилки, а інший – у протилежному. Синтезовані фрагменти Оказакі на 5'-кінці дезоксирибози містять ковалентно зв'язану РНК-затравку. Остання за участю ДНК-полімерази І, яка виявляє ендонуклеазну активність, видаляється. В міру видалення рибонуклеозидних мономерів праймера проходить їх заміна на комплементарні дезоксирибонуклеозидмонофосфати. Здійснюється цей процес за участю ДНК-полімерази І. Далі фрагменти Оказакі під каталітичним впливом ферменту лігази сполучаються між собою й утворюють дочірній ланцюг ДНК. Два новосинтезованих ланцюги, сполучаючись із своїми комплементарними матрицями, утворюють дві дочірні молекули ДНК, кожна з яких містить один материнський і один дочірній ланцюги. Термінація. Механізм термінації, або завершення синтезу ДНК, вивчено недостатньо. В геномі деяких бактерій термінацію здійснює спеціальний термінатор, в інших такий термінатор відсутній. Тому припускають, що реплікація завершується після зустрічі двох реплікативних вилок, які рухаються назустріч одна одній. Процес реплікації проходить з досить високою швидкістю: 1000 – 2000 нуклеотидів за секунду у прокаріот і близько 100 нуклеотидів у еукаріот. Для цього процесу характерна велика точність. Так, у процесі реплікації на 1010 пар нуклеотидів трапляється лише одна помилка. Помилково включений нуклеотид (наприклад, урацил замість тиміну) миттєво видаляються із синтезованого полінуклеотидного ланцюга, і на його місце вводиться новий. Забезпечення точності реплікації є одним з важливих етапів передачі спадкової інформації від одного покоління до іншого. Синтез PHK на матриці PHK. Учений М.C. Гершенсон і американський біохімік Г. Тимін встановили, що крім реплікації існує й інший шлях синтезу ДНК, коли матрицею служить молекула PHK. Оскільки передача генетичної інформації між різними видами нуклеїнових кислот (ДНК ® PHK) називається транскрипцією (переписування), то процес, що протікає у протилежному напрямку (PHK ® ДНК), дістав назву зворотної транскрипції. Каталізує процес фермент РНК-залежна ДНК-полімераза, або ревертаза (зворотна транскриптаза). Цей фермент було виділено в 1969 – 1970 pp. із тканин, інфікованих онкогенними вірусами, а також із самих вірусів цієї групи, зокрема вірусу Раушера лейкемії мишей і вірусу саркоми Рауса. Ревертаза активується іонами Mn2+, Mg2+, причому першими ефективніше, ніж другими. Вивчення будови ферменту, виділеного із вірусів мієлобластозу птахів, показало, що він складається з двох білкових субодиниць з молекулярною масою 110 000 і 70 000 і містить два атоми зв'язаного цинку. Для функціонування ферменту ревертази необхідна затравка і матричний ланцюг PHK. Спочатку на одноланцюговій РНК-матриці за участю ферменту ревертази синтезується одноланцюгова ДНК. Внаслідок цього утворюється гібридна PHK – ДНК молекула, яка далі виконує роль матриці для синтезу комплементарного полінуклеотидного ланцюга ДНК. Цей етап каталізує фермент РНК-залежна ДНК-полімераза. На останньому етапі за участю ферменту РНК-ази від PHK відщеплюється молекула ДНК. Далі така ДНК включається в геном інфікованої, клітини і зумовлює переродження її в ракову. Однак зворотну транскриптазу виявлено і в культурах синцитіальних вірусів, які не викликають злоякісного переродження клітин. За певних умов здатність каталізувати синтез ДНК на PHK виявляють і ДНК-полімераза І та еукаріотичні ДНК-полімерази. Відкриття процесу зворотної транскрипції має важливе теоретичне і практичне значення. Якщо раніше вважалось, що передача генетичної інформації в усіх організмів, за винятком РНК-вмісних вірусів, проходить лише від ДНК до ДНК і від ДНК до PHK, то нині встановлено, що вона може здійснюватись і від PHK до ДНК. Виявлення ферменту ревертази має важливе значення для ранньої діагностики онкологічних захворювань. Ревертаза становить значний інтерес і для генної інженерії (синтез окремих генів). Біосинтез PHK на матриці ДНК. На матриці ДНК синтезується переважна більшість PHK тварин, бактерій і ДНК-вмісних вірусів. Цей процес називають транскрипцією. Він здійснюється за принципом комплементарності азотистих основ нуклеотидів. Інакше кажучи, синтезується ланцюг PHK, нуклеотидна послідовність якого комплементарна послідовності нуклеотидів одного з ланцюгів ДНК. Відмінність полягає лише в тому, що замість азотистої основи тиміну, характерної для ДНК, у PHK включається урацил. Шляхом транскрипції утворюються всі основні види рибонуклеїнових кислот. Це, по-перше, інформаційні або матричні PHK, які надходять у рибосоми і реалізують одержану інформацію в процесі синтезу білка. Інформаційна PHK може нести інформацію про структуру одного, двох і більшої кількості полінуклеотидних ланцюгів. У першому випадку таку іРНК називають моноцистронною, а в другому – поліцистронною. Моноцистронні іРНК характерні для еукаріотів, а поліцистронні – для прокаріотів. Крім іРНК у процесі транскрипції утворюються рРНК і тРНК, які також відіграють важливу роль у біосинтезі білка. Для синтезу PHK на матриці ДНК необхідна наявність усіх чотирьох видів нуклеотидів у формі трифосфатів. Цей процес можна подати схемою:
Реакцію каталізує фермент ДНК-залежна РНК-полімераза або просто РНК-полімераза. Її виявлено в усіх прокаріотичних і еукаріотичних організмах. Найкраще вивчено РНК-полімеразу, виділену з кишкової палички. Вважають, що в кишковій паличці є лише один вид полімерази, яка каталізує процеси синтезу всіх трьох видів PHK. Полімераза – це складний білковин комплекс із загальною молекулярною масою 480000. До складу цього ферменту входить 5 білкових субодиниць: дві a-субодиниці, b-, b¢- і s-субодиниці. Комплекс, який містить дві a-, b-, b¢-субодиниці, носить назву корферменту (від англ. core – серцевина). Він виявляє певну каталітичну активність, однак не здатний вибирати точку (місце) початку транскрипції. Взаємодія корферменту з s-субодиницею приводить до утворення холоензиму РНК-полімерази, який має всі її властивості. Всі субодиниці у холоферменті сполучені між собою слабкими хімічними зв'язками. Характерною особливістю РНК-полімерази, по-перше, є те, що вона за допомогою s-субодиниці здатна вибирати ланцюг ДНК, з якого здійснюватиметься транскрипція, а також місце, з якого розпочинатиметься цей процес. По-друге, для початку синтезу PHK не потребує затравки, як ДНК-полімераза. Слід підкреслити, що РНК-полімераза не просто синтезує PHK, а й регулює цей процес за участю ряду додаткових кофакторів. Тому повністю оправдано застосування до PHK-полімерази поняття „РНК-полімеразна система”. РНК-полімерази виділено також і з культури клітин людини, тимусу телят, печінки щурів. У кожному випадку з ядерної фракції було виділено три різні ферменти: РНК-полімеразу І (A), РНК-полімеразу II (В) і РНК-полімеразу III (C), які відрізнялися між собою кількістю субодиниць у молекулі і їх молекулярною масою. Вважають, що РНК-полімераза І каталізує в основному синтез рибосомальних PHK, РНК-полімераза II – синтез матричних PHK, a PHK-полімераза III, очевидно, – синтез транспортних PHK і 5S рРНК. РНК-полімерази мають два активних центри. Один з них зв'язує 3'–ОН групи рибози кінцевого нуклеотиду ланцюга PHK, який синтезується, і має назву Т-центр, а другий – нуклеотиди, які приєднуються до ланцюга PHK, що росте, і має назву П-центр. Процес матричного синтезу PHK за участю РНК-полімераз поділяють на три стадії: ініціація синтезу, елонгація (подовження) нуклеотидного ланцюга і термінація – завершення синтезу PHK і вивільнення ферменту. В цілому всі ці стадії становлять цикл транскрипції. Ініціація. На цій стадії РНК-полімераза взаємодіє з невеликою ділянкою ДНК, утворюючи комплекс, який здатний розпочинати зв'язування рибонуклеозидтрифосфатів і ініціювати синтез PHK. Хоч РНК-полімераза виявляє велику спорідненість до всіх ділянок ДНК, проте найміцніше вона сполучається лише ділянками, які характеризуються певними послідовностями нуклеотидів на одному з ланцюгів матричної ДНК. Ці ділянки називають промоторами. Вони багаті АТ-парами нуклеотидів і піримідиновими блоками. Вважають, що специфічність взаємодії PHK-полімерази з певними ділянками ДНК-матриці зумовлена наявністю в цих ферментах d-субодиниць (d-факторів). Зв'язування РНК-полімерази з промоторною ділянкою ДНК приводить до часткового розходження спіралі ДНК, що важливо для початку транскрипції PHK. На цій стадії РНК-полімераза за участю Т-центру взаємодіє з 3–ОН групою рибози першого нуклеозидтрифосфату, з якого розпочинається синтез нової молекули рибонуклеїнової кислоти. При цьому від нього не відщеплюється залишок пірофосфату. Першим нуклеотидтрифосфатом в основному є АТФ або ГТФ. Далі РНК-полімераза другим активним центром (П-центром) взаємодіє з другим нуклеотидтрифосфатом. Останній за участю фосфодіефірного зв'язку приєднується до 3'-вуглецевого атома рибози першого нуклеозидтрифосфату. При цьому вивільнюється Т-центр ферменту і відщеплюється пірофосфат від другого нуклеозидтрифосфату. Елонгація. На стадії елонгації перший активний центр ферменту зв'язується з 3¢–ОН групою рибози другого нуклеотиду і РНК-полімераза переміщується (транслокується) на поверхні ДНК в напрямку росту ланцюга. За цих умов другий активний центр ферменту сполучається з наступним (третім) нуклеотидом. Останній знову за участю фосфодіефірного зв'язку сполучається з другим нуклеозидмонофосфатом. Так відбувається ріст (синтез) нуклеотидного ланцюга в напрямку 5 ® 3 до повного завершення біосинтезу PHK. Спід зазначити, що в системах in vivo швидкість елонгації досягає близько 40 – 50 нуклеотидів за секунду, у системах in vitro цей процес проходить менш інтенсивно – 15 – 40 нуклеотидів за секунду. Разом з цим транскрипція здійснюється з досить великою точністю. Частота помилок становить 2×10-3 – 2×10-4 тобто на 2000 – 20 000 нуклеотидів може включатись один неправильний нуклеотид. Хоч ці помилки і малі, однак вони значно більші, ніж помилки при реплікації ДНК, де частота помилок становить 10-10. Однією з причин цього є те, що в процесі реплікації бере участь додатковий механізм контролю – виправляння помилок за участю ДНК-полімерази І. Термінація. Процес обриву ланцюга, порівняно з іншими процесами синтезу PHK менш вивчений. Вважають, що ДНК має специфічні, термінуючі послідовності нуклеотидів. РНК-Полімераза, підходячи до них, перестає функціонувати. При цьому синтезований полінуклеотидний ланцюг PHK і молекула ферменту відділяються від ДНК-матриці. Безпосереднім продуктом транскрипції є різні види PHK. Їх називають первинними транскриптами, або попередниками відповідних цитоплазматичних PHK, і позначають пре-РНК. Процес перетворення даних пре-РНК у функціонально активні форми PHK називається процесингом, або дозріванням. Процесинг відбувається в ядрі і має свої особливості для кожного виду PHK. Важливою групою первинних транскриптів в ядрі є так звана гетерогенна ядерна PHK (гя-РНК). Основну фракцію гя-РНК становить матрична (інформаційна) PHK (пре-мРНК). Для більшості пре-мРНК, синтезованих в ядрі еукаріот, існує три основні етапи процесингу: кепування і метилювання 5'-кінця, поліаденілування 3¢-кінця, видалення (вирізання) некодуючих ділянок (інтронів) з полінуклеотидного ланцюга та сполучення (зшивання) кодуючих ділянок (екзонів). Цей процес називається сплайсингом (від англ. Spticing – з'єднувати, зплітати). Для мPHK, які синтезуються в клітинах прокаріот, процесинг не характерний. Модифікація полінуклеотидних кінців пре-мPHK відбувається так. До 5'-кінця пре-мРНК приєднується олігонуклеотид, який називають „кепом”, або „ковпаком”. „Кеп” складається з двох-трьох метильованих нуклеотидів. Кінцевим нуклеотидом „кепа” є 7-мегилгуанозин, сполучений з полінуклеотидним ланцюгом PHK не по типу 5' ® 3', а 5' ® 5'-фосфодіефірним зв'язком. Вважають, що метильований „кеп” захищає мРНК від руйнування ферментами. До 3'-кінця пре-мРНК приєднується поліаденіловий фрагмент – полі (А), який складається з 150 – 200 аденілових нуклеотидів. Вважають, що полі (А) – фрагмент, очевидно, необхідний для транспорту мРНК з ядра в цитоплазму, а також для підвищення її стабільності. Сплайсинг пре-мРНК відбувається шляхом вилучення неінформативних ділянок (інтронів), починаючи з 5¢-кінця за участю екзо- та ендонуклеаз. Інформативні ділянки, які залишились, тобто екзони, за участю спеціальних РНК-лігаз сполучаються в єдиний полінуклеотидний ланцюг. Певним змінам (процесингу) піддаються й інші пре-РНК, зокрема пре-рРНК і пре-тРНК. Пре-рРНК є попередником рибосомних PHK як у клітинах прокаріот, так і еукаріот. У прокаріот всі три рРНК – 25S, 16S і 5S утворюються з однієї молекули пре-рРНК – 30S, яка має молекулярну масу ~ 2×106. Спочатку окремі азотисті основи, що входять до її складу, піддаються етилюванню. На наступному етапі пре-рРНК розщеплюється з утворенням 17S- і 25S-проміжних PHK. Далі від них поступово за участю нуклеаз відщеплюються нуклеотиди й утворюються характерні для прокаріот 16S і 23S-рРНК. 5S-рРНК утворюється окремо з 3'-кінцевої ділянки пре-рРНК. В еукаріот 18S, 28S і 5,8S-рРНК утворюються в декілька етапів з великої 45S пре-рРНК. Процесинг відбувається в ядерці. Спочатку проходить також метилювання нуклеотидів, а потім відщеплення нуклеотидів, що призводить до утворення 18S, 28S і 5,8S-pPHK. тРНК також утворюються з довших полінуклеотидних ланцюгів пре-тРНК шляхом ферментативного відщеплення зайвих нуклеотидів з 5'- і 3'-кінців молекули. У багатьох випадках з однієї молекули пре-тРНК утворюється дві і більше (до 7) молекул тРНК. У ході процесингу крім відщеплення нуклеотидів з кінців полінуклеотидного ланцюга проходить ще ряд змін. По-перше, з окремих пре-тРНК до 3'-кінця приєднується нуклеотидна послідовність ЦЦА, в інших пре-тРНК цей 3'-кінцевий триплет утворюється в процесі транскрипції. По-друге, багато азотистих основ нуклеотидів зазнають різних модифікацій (деякі метилюються, інші – дезамінуються, відновлюються тощо). І, нарешті, слід зазначити, що синтез PHK істотно відрізняється від реплікації ДHK. Насамперед, він є консервативним, а не напівконсервативним – продукт синтезу не включає жодних компонентів матриці. Цікавим є і той факт, що ферменти синтезу PHK, тобто PHK-полімерази, специфічні відносно різних матриць і навіть їх окремих ділянок. Крім того, для матричного синтезу PHK не потрібна затравка. Процес синтезу починається взаємодією двох нуклеотидів. І, нарешті, процес транскрипції, на відміну від реплікації, проходить переважно з одного полінуклеотидного ланцюга ДНК-матриці.
Розпад нуклеїнових кислот в тканинах
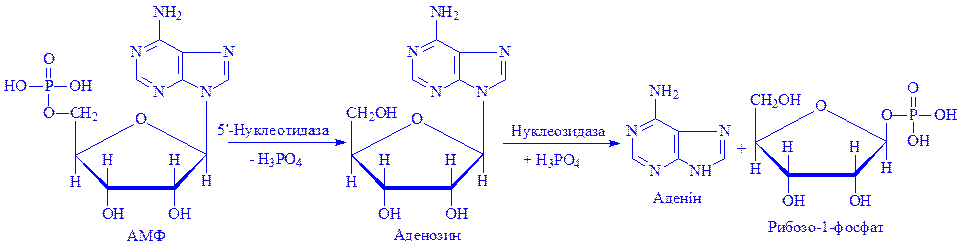
В процесі життя відбувається постійне самооновлення клітин організму. Ці явища пов'язані з синтезом і розпадом молекул ДНК і РНК. Клітини епітелію дванадцятипалої кишки оновлюються протягом 1,6 діб, рогівки ока – 6,9, мезотелія плеври – 33,9, миготливого епітелію трахеї – 47,6 діб. Тривалість життя тромбоцита до 8 діб, лейкоцита до 13, еритроцита до 120 діб. Нуклеїнові кислоти клітин під впливом нуклеаз (ДНК-ази і РНК-ази) розщеплюються до мононуклеотидів. Екзонуклеази розщеплюють до мононуклеотидів зовнішні ділянки молекул ДНК і РНК, ендонуклеази – внутрішні. Частина мононуклеотидів може використовуватися новою клітиною, для синтезу власних нуклеїнових кислот, інші мононуклеотиди розщеплюються на нуклеозид і фосфат, потім – на пентозофосфат і пуринову або піримідинову основу:
На другому етапі розщеплення реакцію каталізують ферменти, специфічні для кожного нуклеозида. Таким чином, у результаті розщеплення нуклеотидів утворюються пуринові і піримідинові основи, фосфати пентоз, фосфорна кислота. Пентози можуть використовуватися як енергетичний матеріал. Основи зазнають ряд глибоких перетворень.
|
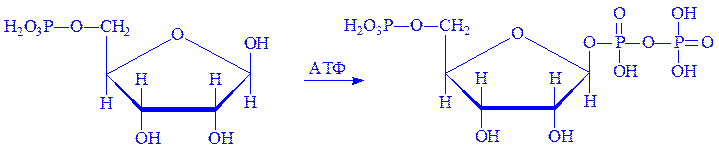
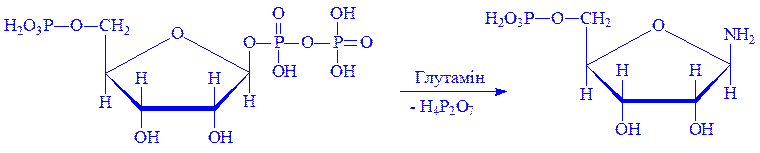

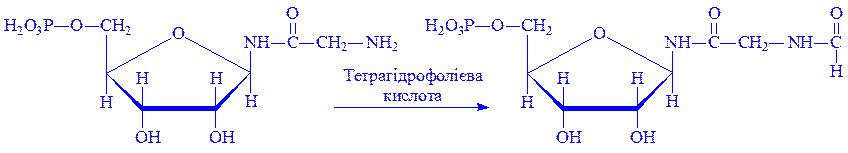




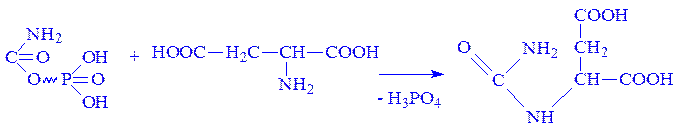


 дТТФ.
дТТФ.