Генетичні засади соціальної поведінки бактерій.
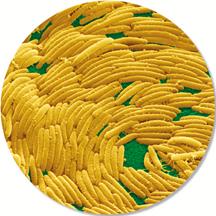
Зазвичай під поведінкою розуміють реакції, що проявляються в рухах. Поведінка бактерій, звичайно, не може бути складною та різноманітною. Але багато бактерій здатні до направленого пересування відповідно до сигналів, що надходять із навколишнього середовища. Є кілька принципово різних типів руху бактерій. Найпоширеніший рух – за допомогою джгутиків: одиночних бактерій і бактеріальних асоціацій (роїння). Окремим випадком цього також є рух спірохет, які ізвиваються завдяки аксіальним ниткам, близьким за будовою до джгутиків, але розташованим у периплазмі. Іншим типом руху є ковзання бактерій, що не мають джгутиків, по поверхні твердих середовищ. Його механізм поки недостатньо вивчений, передбачається участь у ньому виділення слизу (проштовхування клітини) і фібрилярних ниток, що перебувають у клітинній стінці та викликають «хвилю, що біжить» по поверхні клітини. Нарешті, бактерії можуть спливати й поринати в рідині, міняючи свою щільність, наповнюючи газами або спустошуючи аеросоми. У бактерій можна вивчати деякі найпростіші елементи поведінкових реакцій, наприклад, рух, обумовлений мерехтінням джгутиків або війок. Бактерії відповідають на хімічне подразнення рухом у напрямку до джерела подразнення (позитивний хемотаксис) або, навпаки, від цього подразника (негативний хемотаксис). Ця їхня реакція обумовлена функціонуванням своєрідного молекулярного мотора, від якого залежать обертання джгутика, який діє на зразок пропелера й направляє рух бактерії. Бактерії взаємодіють одна з одною, як усередині одного виду, так і із представниками інших видів. При цьому вони обмінюються сигнальними молекулами. Хоча генетика мікроорганізмів вивчається досить активно (повністю розшифровані геноми багатьох сотень мікробів), генетичні основи їхньої поведінки вивчені набагато гірше. Але, проте, є цікаві результати. Так, учені з Інституту біології розвитку ім. Макса Планка (Тюбінген, Німеччина) на прикладі ґрунтової бактерії Myxococcus xanthus показали, що радикальні зміни колективної поведінки та міжорганізмових взаємозв'язків у бактерій можуть відбуватися в результаті досить незначних модифікацій генома. Myxococcus – типовий представник міксобактерій, яким властива досить складна колективна поведінка. Наприклад, вони іноді збираються у великі скупчення та влаштовують колективне «полювання» на інших мікробів. «Мисливці» виділяють токсини, що вбивають «здобич», а потім використовують органічні речовини, що вивільнилися при розпаді загиблих клітин. Як і інші міксобактерії, Myxococcus при нестачі їжі утворює плодові тіла, у яких частина бактерій перетворюється на спори. У вигляді спор мікроби можуть пережити несприятливі умови. Плодове тіло збирається з величезної кількості індивідуальних бактеріальних клітин. Створення такої великої й складної багатоклітинної структури вимагає спільних скоординованих дій мільйонів окремих бактерій, з яких лише мала частина одержує пряму вигоду, а всі інші, по суті справи, жертвують собою заради «загального добра». Тобто далеко не всі з учасників колективної дії зможуть перетворитися на спори й передати свої гени наступним поколінням. Всі інші виступають у ролі «будматеріалу», що загине, не залишивши потомства.
Рис.3.1. Myxococcus - типовий представник міксобактерій
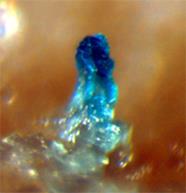
Рис.3.2. Плодові тіла, утворені бактеріями Myxococcus xanthus
Серед міксококів є генетичні лінії (або штами), які не здатні до утворення власних плодових тіл, але вони вміють пристроюватися до «чужих» плодових тіл і, більш того, утворювати там свої спори. Один з таких штамів (дослідники назвали його OC, від obligate cheater – «облігатний ошуканець») не здатен утворювати спори в монокультурі, однак, він успішно проникає в чужі плодові тіла й спорулює там, навіть, із більшою ефективністю, ніж штам-хазяїн, що побудував плодове тіло. Учені експериментували із цією системою «паразит-хазяїн», вирощуючи змішану культуру альтруїстів і ошуканців поперемінно на «голодному» та багатому живильними речовинами середовищі та стежили за коливаннями чисельності двох штамів. Під час голодувань вижити могли тільки ті бактерії, яким удалося перетворитися на спори. В одному з дослідів у штамі OC відбулася мутація, у результаті якої бактерії не тільки відновили втрачену здатність до самостійного (без «допомоги» іншого штаму) утворення плодових тіл і спор, але й одержали додаткову перевагу. По не цілком ясних причинах мутантні бактерії виявилися захищеними від «нахлібників» – представників штаму ОС, своїх прямих предків. Виниклий у результаті мутації новий штам одержав умовну назву PX (від phoenix – «фенікс»). Цей штам з'явився та швидко розмножився саме у той момент, коли майже всі бактерії в експериментальній культурі (вихідний «дикий» штам і паразити ОС) загинули. Штам PX, як з'ясувалося, перемагає в конкурентному змаганні і «диких» міксококів, і паразитів ОС. Він не має потреби для утворення плодових тіл в інших штамах і захищений від паразитизму. Генетичний аналіз досліджених штамів показав, що PX відрізняється від свого прямого предка OC єдиною мутацією – заміною нуклеотида в ділянці гена ферменту ацетилтрансферази, яка нічого не кодує (імовірно є регуляторною). Усього в геномі міксокока більше 30 генів різних ацетилтрансфераз (так називаються ферменти, що прикріплюють до яких-небудь молекул ацетильну групу -CO-CH3), і конкретна функція тієї з них, ген якої змінився в штамі PX, невідома, але деякі непрямі дані, однак, указують на можливу регуляторну роль цього ферменту. Не виключено, що він впливає на активність якихось транскрипційних факторів – білків, що регулюють активність генів. Дійсно, активність цілого ряду генів в PX сильно змінилася в порівнянні з OC. Отже, ученим вдалося зареєструвати мутацію, у результаті якої бактерії Myxococcus xanthus надбали одразу дві корисні властивості: здатність до складної колективної поведінки (утворення плодових тіл) і захищеність від паразитів. Мутація полягала в заміні одного нуклеотида у регуляторній області гена, який, імовірно, бере учать у регуляції активності інших генів. Інша група вчених – з Гарвардського університету – в результаті багаторічних досліджень з'ясувала генетичну основу складної соціальної поведінки Bacillus subtilis. Ці широко розповсюджені ґрунтові бактерії відносяться до числа найбільш добре вивчених мікроорганізмів, їх геном повністю розшифрований в 1997 році. Розмір генома – 4 214 630 пар основ, загальна кількість генів – 4225, і функції більшості генів загалом відомі. Цього, однак, недостатньо, щоб зрозуміти механізми, що керують складною поведінкою цієї бактерії. B. subtilis уміє, наприклад, при необхідності відрощувати джгутики та здобувати рухливість, збиратися в «зграї» (swarming behavior) і приймати рішення на основі хімічних сигналів, одержуваних від родичів. При цьому використовується особливе «почуття кворуму» (quorum sensing) – щось на зразок хімічного голосування, коли певне критичне число поданих родичами хімічних «голосів» змінює поведінку бактерій. B. subtilis здатна збиратися в багатоклітинні агрегати, які за складністю своєї структури наближаються до багатоклітинного організму.
Рис. 3.3. «Плодове тіло» Bacillus subtilis – складний багатоклітинний агрегат, часто утворюваний дикими бактеріями в природі, і майже ніколи – у лабораторії.
У критичній ситуації (наприклад, при тривалому голодуванні) бактерії перетворюються на спори, стійкі до несприятливих впливів. Але перетворення на спору для B. subtilis – процес дорогий та потребує активізації близько 500 генів, і ця міра приберігається на крайній випадок. У якості передостанньої міри в голодні часи B. subtilis прибігає до вбивства своїх родичів і канібалізму. При голодуванні в B. subtilis спрацьовує особливий генний перемикач, що може перебувати лише в одному із двох дискретних станів (включене/виключено). «Перемикач» складається із ключового гена-регулятора Spo0A і декількох інших генів, які взаємно активують один одного за принципом позитивного зворотного зв'язку. Активізація Spo0A призводить до цілого каскаду реакцій, у тому числі до виробництва клітиною токсину SdpC, що вбиває тих бактерій, у яких «перемикач» вимкнений. Але голодування призводить до активізації Spo0A тільки в половини мікробів. Загиблі клітини розпадаються, а органічні речовини, що вивільнилися з них, використовуються «вбивцями». Якщо ніяких змін на краще так і не відбудеться, вони таки утворять спори. Спочатку було неясно, чому токсин убиває тільки тих бактерій, які його не виділяють (тобто тих, у кого Spo0A не активований). Виявилося, що на мембрані бактерій є захисний білок SdpI, що виконує одразу дві функції. По-перше, він захищає клітину від токсину SdpC (просто хапає молекули токсину і тримає, не дає їм нічого робити). По-друге, молекула білка SdpI, що схопила молекулу токсину, змінюється таким чином, що інший її кінець (той, що стирчить із внутрішньої сторони мембрани) хапає та утримує молекули білка SdpR, функція якого полягає в тому, щоб блокувати виробництво захисного білка SdpI. Таким чином, захоплення захисним білком молекули токсину призводить до інактивації білка, що гальмує виробництво захисного білка. Тобто чим більше буде токсину, тим більше клітина буде робити захисного білка. А як тільки токсин у навколишнім середовищі скінчиться, молекули SdpR припинять інактивуватися, і виробництво захисного білка зупиниться. За молекулярно-біологічними мірками це – вкрай простий регуляторний каскад. Так бактерії захищаються від власного токсину. А бактерії з вимкненим Spo0A виявляються незахищеними, тому що синтез рятівного SdpI у них блокується ще одним білком – AbrB. Вимкнути AbrB можна тільки шляхом включення Spo0A, тому клітини з вимкненим Spo0A приречені на загибель.
|