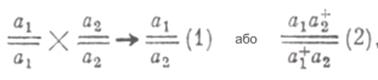Генетичний аналіз поведінки за допомогою спонтанних і індукованих мутацій.
Для вивчення поведінкових ознак важливо одержати досить представницькі вибірки мутацій, що на них впливають. При наявності таких вибірок стає можливим виявити участь різних генів у контролюванні даної ознаки, розкрити характер їхньої взаємодії, підійти до механізмів генетичного регулювання поведінкових особливостей. Питання про необхідність одержання в достатніх кількостях потрібних поведінкових мутацій у дрозофіли був поставлений С. Бензером у сімдесятих роках ХХ століття. Бензер вважав, що завдання зводиться до створення аналогів селективних середовищ, де в якості фактора селекції виступала б сама поведінкова особливість мутантних мух. У відповідь на відповідний подразник вони повинні автоматично відділятися від іншої маси нормальних особин. Надійність виділення потрібних мутантів підвищують за рахунок багаторазового повторення процедури сортування особин за поведінкою. Свої ідеї Бензер реалізував при одержанні мутацій, що впливають на фотореакцію, при індукції їх хімічними мутагенами. Використовували сильний алкілуючий агент етилметансульфонат, що викликає у великій кількості точкові мутації типу заміни пар основ, та методику «Muller-5», яка зазвичай застосовується для виділення рецесивних зчеплених зі статтю мутацій, а також деякі інші методики й лінії зі зчепленими А-хромосомами в самиць.
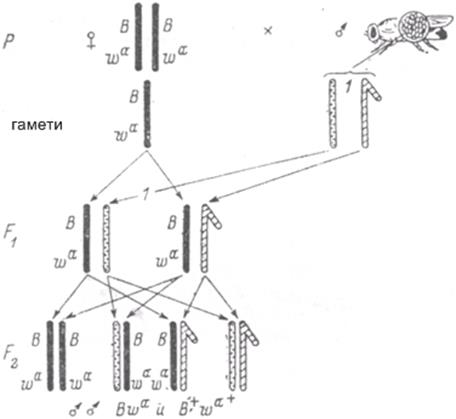
Рис. 1.39. Метод виявлення рецесивних зчеплених зі статтю летальних мутацій у дрозофіли (методика «Muller-5») В – ген смугопобіних очей; wa – ген абрикосового кольору очей.
Ефективний спосіб виділення термочутливих неврологічних мутантів розробив в 1971 році Сузуки. Він, так само як і Бензер, використовував для індукції мутацій хімічні мутагени. Високоефективне селективне середовище для добору фізіологічних мутантів з нульовою активністю алкогольдегідрогенази (АДГ) розробили в 1972 році Софєр і Хеткоф. Таким чином, завдання зі створення селективних методик для виділення фізіологічних і поведінкових мутацій у дрозофіли було вирішене. Але одержання мутацій за певною ознакою є лише першим етапом генетичної роботи. Другий етап вивчення генетики поведінки за допомогою індукованих мутацій полягає у встановленні генів, до яких належать виділені мутації. Імовірні три ситуації: · всі виділені по даній ознаці мутації будуть результатом мутування одного гена; · кожна окрема мутація буде результатом змін окремого локусу; · мутує кілька локусів, причому деякі з них неодноразово. Для визначення приналежності двох рецесивних мутацій до того самого чи різних генів використовують функціональний тест на алелізм. Для цього їх поєднують у гетерозиготному стані в потомстві від схрещування гомозиготних батьків:
де а1 і a2 – мутації за даною поведінковою ознакою.
Якщо в гетерозиготних особин F1 виявиться аномальна поведінка, то можна вважати, що аналізовані мутації не є комплементарними одна одній, тобто є алелями одного гена, і їхній генотип варто записувати як (1). Якщо ж у гетерозигот F1 виявиться нормальна поведінка, значить в аналіз залучені комплементарні, взаємодоповнюючі мутації, які відносяться до різних локусів (2). У роботах Хотта, Бензера, Сузукі було випробувано на алелізм по кілька десятків рецесивних зчеплених зі статтю мутацій. Мутації, що змінюють фототаксис, виникли, як виявилося, у результаті мутування семи різних генів у Х-хромосомі, а температурозалежні мутації, що викликають параліч, розподілилися між чотирма генами. Третій етап генетичного аналізу полягає в картуванні локусів, по яких розподілені виділені мутації. Картування нових мутантних генів, у тому числі й тих, що впливають на поведінку, досягається за допомогою ліній-тестерів, що несуть у відповідній хромосомі низку рецесивних і домінантних маркерів, таких ознак, які легко відрізнити та гени яких точно локалізовані. У гібридному потомстві враховують частоту рекомбінантних класів особин, що несуть досліджувану ознаку й маркерні ознаки. Необхідний результат може бути також досягнутий за допомогою тестерних ліній, що несуть серію делецій уздовж хромосоми. Гени, що контролюють дану поведінкову ознаку, можуть розподілитися на генетичній карті будь-яким образом. Наприклад, Каплан зі співробітниками досліджував мутації, що порушують локомоцію й чітко проявляються на провокаційному тлі, – при ефірному наркозі. Тридцять отриманих ними мутацій розподілилися по чотирьох локусах у Х-хромосомі на ділянці близько 30% довжини генетичної карти, тобто в 30 морганід. В іншій роботі 53 термочутливі мутації з 52 локусів, зосередилися на одній ділянці, що займає 13% довжини генетичної карти 3-ї хромосоми. Доведення генетичного аналізу до картування окремих генів, що контролюють фізіологічну або поведінкову ознаку, дозволяє зрозуміти генетичну організацію цієї ознаки. Фенотипічну локалізацію дії мутантного гена можна здійснити за допомогою хромосомних мозаїків. У сімдесяті роки ХХ століття вченими Хотта й Бензером були отримані мозаїчні особини, що несуть різні по величині нормальні й мутантні частини тіла. В експерименті використовувалися раніше отримані рецесивні мутації, які розподілені по п'яти локусах Х-хромосоми та порушують фототаксис. Ці мутації шляхом кросинговеру нарізно сполучалися в одній хромосомі з рецесивними маркерами: у w sn f. Гомологічна Х-хромосома несла нормальні алелі цих рецесивних мутацій, але включала структурну перебудову, що надавала їй форму кільця. Така незвичайна кільцева хромосома вже на ранніх ембріональних стадіях розвитку має високу ймовірність перетерпіти розриви та, у результаті, втратитися. Ті тканини, у клітинах яких збережуться обидві Х-хромосоми, будуть за фенотипом нормальними. Ті ж тканини й органи, які розів'ються з клітин, що втратили кільцеву Х-хромосому, виявляться мутантними, оскільки рецесивні мутації, будучи в гемізиготному стані, зможуть тепер виявитися. Чим раніше в онтогенезі відбудеться елімінація кільцевої Х-хромосоми, тим більша ділянка тіла буде мати мутантний фенотип. При наявності маркерів, що змінюють забарвлення тіла (у), ока (w) і форму щетинок (sn і f), мутантні тканини легко розрізнити на поверхні будь-якої ділянки тіла.
Рис. 1.40. Гінандроморф дрозофіли: лівий бік тіла (чорний) складається із клітин із двома Х-хромосомами, а правий (білий) – з клітин з однією Х-хромосомою.
Рис. 1.41. Інші варіанти будови гінандроморфів дрозофіли. Типи мозаїчності фенотипу в гінандроморфних особин D.melanogaster по мутації hyperkinetic.
Відомо, що в складному оці дрозофіли є близько 800 оматідієв. (Оматідій – це структурна й функціональна одиниця фасеточного ока комах. Оматідії сформувалися в ході еволюції з ізольованих простих вічок при їхній інтеграції в складні, або фасеточні, очі). Кожний оматідій включає 8 впорядковано розташованих фоторецепторних клітин. Хотта й Бензер знімали електроретинограми – ЭРГ – (це запис біоелектричних потенціалів сітківки ока, що виникають при дії на неї світла) у мозаїчних особин, що мали одне око нормальне, а інше – мутантне, або ж частину ока нормальну, а іншу – мутантну. В окремих випадках досліджували мозаїків з порушенням лише кількох оматідіев. У результаті був доведений високий ступінь автономності фоторецепторних клітин ока, функціонування яких визначається особливостями їх власного генотипу. Одночасно з роботою Хотта й Бензера з'явилася робота Ікєди та Каплана, які застосували метод одержання мозаїків для нейрофізіологічного дослідження мутацій, що викликають посмикування лапок у наркотизованих ефіром мух. Було вивчено 77 гінандроморфів, що несуть у Х-хромосомі відповідну мутацію. За допомогою мікроелектродної техніки одержали характеристики електричної активності окремих нейронів торакального ганглія й довели автономність окремих мотонейронів у про-, мезо- і метаторакальних відділах торакального синганглія. Без використання генетичних підходів одержання таких результатів навряд чи було б можливим. Дослідження мозаїчних особин дозволило Хотта й Бензеру запропонувати спосіб вичленовування так званих фокусів - структур, відповідальних за прояв поведінкових ознак. Логіка міркувань при цьому була такою. У ранньому ембріогенезі дрозофіли, вже на стадії бластодерми, окремі ділянки ембріону детерміновані. З них формуються імагінальні диски, які, у свою чергу, дають початок певним частинам тіла імаго. Якщо мозаїцизм захоплює дану ділянку тіла, що виявляється за допомогою маркерів, і це супроводжується певною поведінковою аномалією, то тим самим доводиться ураження нервової системи, мускулатури або інших структур саме в даній ділянці. Таким шляхом було підтверджено, що мутації, які порушують зорову рецепцію та змінюють електроретинограму, порушують клітини ока або ділянки, що безпосередньо до них примикають. Мутація Hk1P порушує функцію торакального ганглія. Складний фокус мутації drop-dead (drd), що викликає ранню загибель мух, перебуває в церебральному ганглії. Білатеральні взаємозалежні фокуси мутацій wings-up локалізуються в районі торакса. Одержання мозаїків можливе лише при втраті однієї з Х-хромосом. Якщо ж втрачається одна з великих аутосом, тоді дуже швидко наступає загибель особини. Але Холл і Кенкєль, співробітники Бензера, транслокували на Х-хромосому невеликі ділянки аутосом, нестача яких у гетерозиготному стані не призводить до летального результату. Ці дослідники у якості маркерів використовували мутантні гени кислої фосфатази та α-гліцерофосфатдегідрогенази, що дало можливість спостерігати мозаїцизм тканин також і на внутрішніх органах. При вивченні генетики поведінки використовують і неврологічні мутації. Наприклад, Лєвін і Вімен провели дослідження морфології літальних м'язів і їхньої іннервації, використовуючи для цього мутацію stripe. Мутантні гомозиготні мухи не здатні літати, їхня моторна активність редукована до короткочасних високочастотних фібриляцій крил, однак видимих морфологічних відхилень від норми вони не мають. Ґрунтуючись на результатах мікроелектродної техніки, автори дійшли висновку, що ненормальна поведінка мух stripe зумовлена дисбалансом у передачі імпульсів, що направляються синергічним волокнам, і імпульсів, що направляються м'язам-антагоністам. Конопка й Бензер провели дослідження циркадного ритму за допомогою серії спеціально отриманих мутацій. При застосуванні хімічного мутагену етилметансульфонату (ЭМС) були отримані три зчеплені зі статтю рецесивні мутації, що змінюють добову циклічність виходу імаго з пупарієв: per";, per', per1. Всі три мутації локалізовані на короткій ділянці Х-хромосоми між генами zeste (1.0) і white (1.5). Мутація null-period-arhythmic (per0) зумовлює порівняно рівномірний розподіл частот відродження мух протягом доби. Мутація short-period (pers) контролює цілком чіткий ритм, але не при 24-годинному, а при 19-годинному циклі. Мутація long-period (perl) подовжує цикл виходу до 28 годин. У лабораторії Бензера в 1976 році була отримана зчеплена зі статтю мутація dunce («тупиця»). Особливість цих мутантів полягає в нездатності до вироблення умовних рефлексів уникання на специфічний запах при підкріпленні електричним струмом. У той же час їх нюхові реакції, так само як інші фізіологічні й поведінкові особливості, зберігаються нормальними. Отримані мутації, що зумовлюють резистентність до таких нейротропних і психотропних речовин, як амфетамін, барбітал, бідрин, фізостігмін, кофеїн, теофілін і пікротоксин. Ці мутації представляють інтерес для дослідження метаболізму нейромедіаторів. Одержання мутацій, що впливають на фізіологічні й поведінкові ознаки, відкриває сприятливі можливості для їх наступного поглибленого дослідження. Генетичний аналіз таких мутацій може бути здійснений повною мірою, аж до картування на генетичній карті. З їхньою допомогою може бути розкрита система генетичного контролю фізіологічних ознак. Внесок мутаційного аналізу поведінкових ознак у генетику поведінки є досить значним.
|