Генетика морфологічних особливостей нервової системи та їх зв'язок з мінливістю ознак поведінки.
Генетиків завжди цікавила морфофізіологічна основа поведінки. Знання цієї основи – необхідна передумова для успішного пошуку механізмів впливу генів на поведінку. Використання штучно одержуваних мутацій при гарному знанні генетики досліджуваного об'єкта, його розвитку, нейроанатомії допомагає розкрити первинні морфологічні, електрофізіологічні або біохімічні ефекти генів, що ведуть до зміни поведінки. Варіації структурної організації нервової системи також впливають на поведінку. Одним з показників нейронного статусу є вага мозку. На мишах виявлені лінійні відмінності по вазі мозку, але ці відмінності не корелювали, наприклад, з такою ознакою, як швидкість надбання умовних реакцій уникання при дії електричного струму. Однак генетична модифікація ваги мозку селекційним шляхом впливає на здатність до навчання. У процесі селекції змінюються темпи й тривалість розвитку нервової тканини, причому селекція на високу вагу мозку збільшує темпи ембріонального й неонатального росту, але не міняє його тривалості. Тварини цих ліній мають більш важкий мозок при народженні в порівнянні з неселектованим контролем, і ця різниця зберігається протягом усього періоду постнатального росту. Селекція у зворотному напрямку не змінює темпів ембріонального розвитку нервової тканини. Вага мозку в немовлят селектованих тварин однакова у порівнянні з вагою мозку особин з контрольної групи. Однак у перших наступає більш рання зупинка в постнатальному розвитку мозку. Це може мати істотний вплив на поведінку, тому що разом зі скороченням періоду морфологічного розвитку мозку змінюється й тривалість періоду, залежного від середовищної стимуляції функціонального становлення нервової системи. Установлено, що фактори середовища в ранній період онтогенезу істотно впливають на деякі морфологічні показники нервової тканини. Виявилося, що збільшення числа мікронейронів (зірчастих, ретикулярних і гранулярних нервових клітин), яким приділяється особлива роль у здійсненні творчої функції мозку, відбувається й після народження. Вплив певних факторів середовища в ранньому постнатальному критичному періоді розвитку призводить до збільшення числа мікронейронів і до ускладнення їхніх дендритних зв'язків. Тому селекція на більш низьку вагу мозку, скорочуючи період його постнатального розвитку, може зменшувати як число нейронів, так і число міжнейронних контактів. Селекція на кращу здатність до навчання спричиняє збільшення ваги мозку й особливо одного з його відділів – гіпокампу, що відіграє важливу роль у процесі навчання. Гранулярні нейрони зубчастої фасції гіпокампу мають пряме відношення до зберігання слідів минулого досвіду. Описані міжлінійні відмінності в структурній організації провідних шляхів головного мозку мишей. Так, у ліній, що характеризуються крайньою агресивністю, високою руховою активністю або гарною навченістю, спостерігалися різноманітні відхилення від нормальної організації провідних шляхів, наприклад, деяка редукція мозолистого тіла, зміна положення передньої комісури, і деякі інші відхилення в розташуванні провідних шляхів, які могли бути основою структурної пластичності мозку й причиною спадкової мінливості поведінки. Генетичний аналіз показав, що виявлена мінливість у структурній організації провідних шляхів детермінована полігенно. При аналізі клонів коника К. Гудман порівняв однорідність і варіабельність росту й розвитку різних нейронів. Для багатьох нейронів клонованих коників були показані розходження в утворених нейральних зв'язках. Подібні результати були отримані на мікроскопічних ракоподібних – дафніях. Самиці дафній, як відомо, здатні давати потомство, не будучи заплідненими самцями. Завдяки цій здатності до партеногенезу в даного виду можливо одержати лінії генетично ідентичного потомства однієї самки – клони. Око дафнії містить 176 сенсорних нейронів, які утворюють синаптичні контакти рівно зі 110-ма нейронами оптичного ганглія. Але точне число синапсів, що утворяться між певним сенсорним нейроном і специфічним нейроном ганглія, може відрізнятися більш ніж у три рази при порівнянні окремих клонів. Припускають, що подібна варіабельність обумовлена дією зовнішніх факторів. Набагато складніше виявити ідентичні нейрони для порівняння синаптичних зв'язків у хребетних тварин. Подібне дослідження можна проводити на близнюках. Деякі костисті риби мають ідентифіковані нейрони – маутнерівські клітини, а деякі види цих риб здатні до розмноження без запліднення; тобто мати й усі її доньки генетично ідентичні. Кожна особина має по одній гігантській маутніровській клітині в кожній половині мозку. Відзначено, що патерн розгалуження дендритів маутнерівських нейронів у клонів подібний, але існують індивідуальні відмінності в деталях розгалуження й утворених синапсах.
Рис. 2.1. Імовірна схема роботи гальмівного електричного синапсу на прикладі маутнерівського нейрону риби: 1 - нервове закінчення, 2 - початковий сегмент аксона маутнерівської клітини, 3 - гліальна «чашечка». Стрілкою показаний напрямок струму, породжуваного потенціалом дії нервового закінчення.
Цікавими є результати експериментального збільшення маси мозку в риб. Шляхом трансплантації на ранніх стадіях ембріонального розвитку був експериментально збільшений відповідний корі ссавців відділ мозку (tectum) в африканської рибки (Tilapia macrocephala). Це призвело до помітного поліпшення її інтелектуальних здатностей.
Рис.2.2., Рис.2.2.а. Африканська рибка (Tilapia macrocephala), яка «порозумнішала» у результаті експериментального збільшення одного з відділів мозку
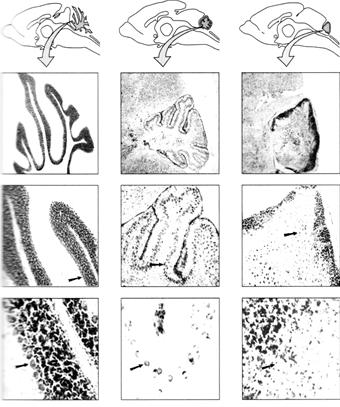
Окремі неврологічні мутації в мишей також викликають ті чи інші структурні зміни в нервовій системі: порушення структури шарів кори головного мозку, клітинних взаємодій у тканині мозочка, ступеня мієлінізації нервових волокон. У мишей відомо багато мутацій, що впливають на нервову систему. У деяких мутантів відсутні певні ділянки мозку. В інших – спостерігаються специфічні анатомічні порушення, такі як дефекти мієлінізації або дезорганізація характерних клітинних утворень, порушення структури шарів кори головного мозку. Одна група мутацій, яка особливо зацікавила дослідників, включала різні моногенні порушення, що впливають на постнатальний розвиток мозочка. Всі вони одержали назви, що чітко характеризують супутні локомоторні порушення: reeler («той, що крутиться»), staggerer («той, що хитається») і weaver («той, що погойдується»).
Рис. 2.3. Мутації, що порушують розвиток мозочка в мишей: а) Мозочок нормальної миші; б) Мозочок мутанта weaver; в) Мозочок мутанта reeler.
Мозочок мутантів reeler характеризується аномальним розташуванням клітин. Спостерігається відсутність характерних шарів клітин у мозочку, гіпокампі чи корі мозку. Мозочок мутантів weaver містить набагато менше гранулоцитів, у порівнянні з нормальним мозочком, внаслідок порушення здатності даного типу клітин нормально мігрувати. Гранулярні клітини мишей weaver не здатні виробляти білок САМ (білок cell adhesion molecule), що забезпечує гранулярним клітинам здатність приєднуватися або мігрувати уздовж гліальних клітин. Рівень гранулярних клітин у мутантів не досягає норми, а в мозочку не формується нормальна ламінарна (пошарова) організація. Р. Грінспан зі співробітниками описав мутантів плодової мушки Drosophila, які були нормальними за всіма ознаками, за винятком того, що мали порушення пам'яті. Ці мутанти, названі dunce («тупиця»), amnesiac («з амнезією») і turnip («ріпа»), або були не здатні до навчання, або могли навчатися, але швидко все забували. Дефекти пам'яті були обумовлені біохімічними змінами, що відбувалися внаслідок інактивації генів і відсутності або порушенні синтезу відповідних білків. Дослідження на дрозофілі виявили велику кількість специфічних мутацій, кожна з яких призводить до певного дефекту в якій-небудь частині нервової системи комахи. Наприклад, летальна мутація Notch викликає в дрозофіли збільшення (розростання) нервової системи внаслідок надлишкового вироблення клітин-попередників.
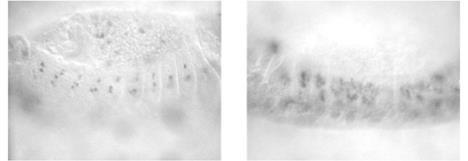
Рис. 2.4. Мутація Notch дрозофіли. В ембріона дикого типу (А) виявляється біля чотирьох клітин попередників нервових клітин на один гемісегмент, а в мутантних (Б) – більше десяти, що є наслідком низької активності гена Notch.
Дослідження генетики морфологічних особливостей нервової системи й зв'язку цих особливостей з мінливістю поведінки активно розвиваються. Експериментальних даних у цій області щодо конкретних зв'язків специфічних форм поведінки зі специфічними структурними особливостями нервової системи поки небагато, однак учені вважають, що морфологічна організація мозку – один з головних шляхів реалізації генетичної інформації на рівні поведінки.
|