БИОСИНТЕЗ ФЛАВОНОИДОВ
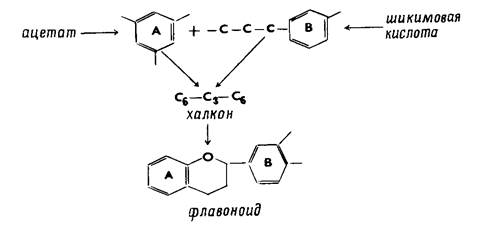
Метаболизм фенилпропаноидных соединений в растениях непосредственно связан не только с биосинтезом лигнина, но и с образованием прочих фенольных соединений, например флавоноидов. Различные группы флавоноидов могут быть произведены от основного звена диарилпропана С6-С3-С6, которое могло бы возникать в растительных органах конденсацией С6 флороглюцина и фрагмента С6 — С3 с пирокатехиновым ядром; впервые эта гипотеза была выдвинута Робинсоном. Оба эти фенола связаны вместе триозой путем альдольной конденсации, причем сначала протекает конденсация пирокатехина с триозой (образование фенилпропанового звена) и лишь затем присоединяется флороглюцин.
Биосинтез флавоноидов по Робинсону
Гипотетический дифенилпропановый промежуточный продукт обеспечивает возможность превращения в различные продукты в результате окисления, дегидратации, восстановления и замыкания пропанового звена в гетероциклическое ядро. Окисление в этом ядре при атоме углерода С-1 приводит к образованию антопианина, при С-3 - к флавонам, при С-1 и С-3 или при С-2 и С-3 - к флавонолам, а при восстановлении - к флаванам. Представления Робинсона можно уточнить в том смысле, что образование флороглюцинового звена (цикл А) происходит по ацетатному механизму биосинтеза фенольных соединений, а пирокатехиновый фрагмент образуется при участии шикимовой кислоты. Из обоих фрагментов могут образоваться халконы - основные соединения типа С6— С3— С6, удобные промежуточные продукты в био- синтезе флавоноидов, согласно схеме
Исследование биосинтеза флавоноидов радиоизотопным методом подтвердило предположение, что звено С6 (цикл А) происходит от ацетата, а звено С6- С3 - от шикимовой кислоты. Было исследовано образование кверцетина, цианидина, флоридизина и катехинов. Весьма интересную работу по биосинтезу катехинов провел Запрометов, который использовал меченый 14Сацетат, шикимовую кислоту и в качестве меченых сахаров глюкозу, фруктозу и сах арозу и изучал образование катехинов в молодых побегах чая. На основании этих экспериментальных данных был предложен механизм биосинтеза катехинов.
Все работы по биосинтезу флавоноидов подтвердили, что флороновое ядро А образуется по ацетатному механизму - конденсацией трех молекул уксусной кислоты. Предполагается участие ацетил кофермента А или малонилкофермента А. Циклизация остатка β-поликетокислоты приводит к образованию не идентифицированных до настоящего времени просежуточных продуктов. Наиболее вероятно, что фенольные гидроксилы в положениях 5 и 7 происходят от карбоксильных групп молекул уксусной кислоты. Вторая часть молекулы флавоноидов - фенилпропановая (цикл В)- происходит от фенилаланина. При радиоизотопных исследованиях биосинтеза флавоноидов всегда обнаруживалось, что шикимовая кислота вступала в молекулу флавоноида. Тем самым обосновывается взгпяд, что биогенетический путь образования этой части молекулы протекает через шикимовую и префеновую кислоты с образованием фенилаланина и производных коричной кислоты, которые по всей вероятности присоединяют триацетатное производное, образуя промежуточный халкое. Гидроксиилирование в цикле В происходит уже на уровне продукта С 9, и следует полагать, что возникает гидроксилированная коричная кислота. Сходство биогенезиса флаванов, антоцианинов позволяет прийти к заключению, что растительные флавоноиды образуются по одинаковому биогенетическому механизму.
|