
Оценка биологического разнообразия
Весьма сложным методом, но колоссально информативным является оценка таксономического разнообразия. Исследователю при использовании упомянутых методов оценки не требуется исследовать колоссальные базы мониторинговых данных по абиотическому компоненту и определять различные смешанные эффекты этих взаимодействий – наличие определённых видов уже говорит само за себя, но данные методы позволяют делать выводы не только по наличию видов-биоиндикаторов, а по совокупности всей компонентов «биологического разнообразия» (биоразнообразия). Понятие «биологическое разнообразие» за весьма короткий отрезок времени получило большое количество толкований. В биологическом смысле рассматриваются представления о внутривидовом, видовом и надвидовом (ценотическом) разнообразии жизни. Однако, позднее, сначала деятели охраны природы, а затем и ученые стали говорить об экосистемном и ландшафтном разнообразии как объектах сохранения, а, соответственно, изучения и выделения в природе. В 1992 г. на Конференции ООН по окружающей среде и развитию была принята «Конвенция о биологическом разнообразии», к которой присоединилось большинство стран на планете, а, следовательно, сложнейшая и многообразнейшая проблема биоразнообразия приобрела еще и политическое звучание. Большинство исследователей сходятся во мнении, что словосочетание «биологическое разнообразие» впервые применил Г. Бэйтс в 1892 г. в работе «Натуралист на Амазонке», когда описывал свои впечатления от встречи около 700 разных видов бабочек за время часовой экскурсии. Однако основные научные концепции биоразнообразия были сформулированы лишь в середине ХХ века, что напрямую связано с развитием количественных методов в биологии. Особенно отметим работы Р. Уиттекера, в которых была предложена организация уровней экосистемного разнообразия и исследованы зависимости биоразнообразия от факторов окружающей среды. Обратим внимание, что исследование различных аспектов биоразнообразия является одним из магистральных направлений концепции устойчивого развития. Устойчивое развитие определяют как управляемый процесс коэволюционного развития природы и общества. Как деятельность, устойчивое развитие нуждается в поддержке мощной информационной базы цифровых данных по экологическим, социально-демографическим, производственно-экономическим условиям и тенденциям их развития. На основе этих данных строятся модели экономического благополучия и проводятся научные исследования. Здесь мы сталкиваемся с рядом проблем. Во-первых, объемы первичной информации устойчивого развития территорий, даже локального уровня, очень велики. В связи с этим проявляется проблема накопления, целенаправленного структурирования и обработки колоссального объема информации. Во-вторых, существует проблема получения качественно новой информации на основе первичной (здесь следует отметить, что на данный момент накопилось большое количество тематической информации в различных форматах) и прогнозирования. Анализ информации невозможен без адекватного математического аппарата и на его разработку в различных областях тратится «львиная доля» ресурсов. В-третьих, необходима единая концептуальная система, объединяющая этапы сбора, обработки, анализа информации и прогноза. В настоящее время существует несколько попыток реализации информационных систем с экологическим наполнением. Как правило, подразумевается реализация с помощью некоторой ЭИС (экологическая информационная система) или БИС (биоинформационная система). Но основанная проблема всех систем – это математическое обоснование. Определим для начала основные показатели биологического мониторинга. Встречаемость (частота встречаемости, коэффициент встречаемости) – это относительное число выборок (участков), в которых встречается вид. Если выборка состоит из 100 учетных площадок, а вид отмечен на 43, то и встречаемость равна 43%. При встречаемости 25%, вид встречается в каждой четвертой площадке учета и он случайный. Высокая встречаемость, если вид отмечен более, чем на 50% учетных площадок. Обычно закладывается 50 учетных площадок, но не менее 25. Обилие – это количество особей вида на единице площади или объема. Наиболее часто используются шкалы обилия Друде и Хульта (табл. 9.2). Покрытие – процент площади, покрываемой надземными частями растений. Процент площади, занятой основаниями растений – истинное покрытие, верхними частями – проективное. Проективное покрытие – обязательный показатель при изучении надпочвенного покрова. При изучении древесно-кустарниковых ярусов синонимом проективного покрытия служит сомкнутость – отношение площади проекций крон к площади занимаемого участка; в отличие от проективного покрытия сомкнутость измеряется в долях от единицы. Истинное покрытие для древостоя – сумма площадей поперечного сечения стволов и полнота, определяется расчетным путем по данным перечета древостоя. Таблица 9.2 Шкала обилия Друде и шкала обилия Хульта (балльная)
Биомасса – общие запасы органического вещества, накопленные к моменту учета. Выражаются в массе абсолютно-сухого, воздушно-сухого или сырого вещества. Биомасса растений – растительная масса, фитомасса; биомасса животных – зоомасса. Биомасса, ее фракционная структура, скорость накопления (продукция – прирост биомассы за определенный промежуток времени) являются важнейшими – интегральными, показателями жизнедеятельности организмов. Они дают возможность оценить роль каждого фактора и популяции в формировании биогеоценоза, оценить запасы биологических и пищевых ресурсов, сделать кратко- и долгосрочные прогнозы развития сообществ, предсказать пути их трансформации и разработать мероприятия по охране и рациональному использованию любого из ресурсов. Именно поэтому изучение биологической продуктивности и было положено в основу упомянутой выше Международной биологической программы (МБП). При экологических исследованиях очень важна и хозяйственная оценка исследуемых территорий: запасы древесины, лекарственного сырья, пищевых и промысловых ресурсов. В целом же часто необходимо сочетание всех перечисленных методов. Существует несколько областей потенциального применения математических методов оценки компонентов биоразнообразия. Во-первых, это охрана окружающей среды, в основе которой лежит мысль о том, что богатые видами сообщества лучше бедных видами. Во-вторых, мониторинг окружающей среды, исходящий из корреляции уровня загрязнения со снижением разнообразия или изменением характера распределения видового обилия. В обоих случаях разнообразие используется как индекс благополучия экосистемы, а, следовательно, очевидным способом получения подобной информации является количественная оценка компонентов биоразнообразия различных уровней организации. И, наконец, в-третьих, исследование структуры сообществ всегда даёт новую информацию об исследуемом объекте и, как следствие, новый импульс для развития науки и корректировки методов практической деятельности. Мы будем рассматривать процесс оценки компонентов биоразнообразия на основе полной схемы сравнительного анализа в рамках схемно-целевого подхода Б.И. Сёмкина [47] с использованием информационных технологий. Несмотря на то, что исследование структуры биологических объектов – одна из центральных проблем многих биологических наук, таких как экология, геоботаника, фитоценология и др., о содержательной концепции до сих пор идут споры. Когда дело касается количественных оценок, привлечение математических приемов неизбежно, особенно если исследователь работает в некой новой, междисциплинарной области, а тем более, если изучается структура таких сложных объектов природы, как растительный покров. «Первой ласточкой» стала работа П. Жаккара в 1901 г. Этим швейцарским исследователем был предложен коэффициент флористической общности, который и по настоящее время не потерял своей актуальности. В дальнейшем, было опубликовано огромное количество теоретических разработок и работ, касающихся применения количественных методов в конкретной предметной области. Особенно преуспели британские математики – Р.Э. Фишер, К. Пирсон и В. Уэлдон, которых считают основателями биометрии. В отечественной биологической науке можно отметить публикации А.А. Любищева (члена Международного биометрического общества), да ещё, пожалуй, работы Р.Л. Берга в области стабилизирующего отбора, а также работами П.В. Терентьева о методе корреляционных плеяд. В 60-70 гг. в направлении количественной биологии можно отметить явный научный бум. Были опубликованы крупные зарубежные и отечественные обзоры количественных методов в экологических исследованиях [9, 16 и др.], а также огромное количество работ посвящённых отдельным аспектам теории, введению и применению мер сходства-различия (работы Б.И. Сёмкина). Здесь хотелось бы также отметить ряд более поздних обзорных работ по количественным методам в биологических науках [32, 40]. Начиная с первых работ по количественным методам и по настоящее время, в различных естественных науках, наблюдается разрыв между схемами сбора данных и схемами последующей математической обработки. Следствием подобной практики является обычно то, что математические методы очень часто используются некорректно. Во избежание подобных ошибок Б.И. Сёмкиным была разработана концепция схемно-целевого подхода. В общем можно отметить, что суть схемно-целевого подхода заключается в использовании строгой схемы исследования, начиная с выбора цели исследования и заканчивая интерпретацией данных. Рассмотрим данную схему подробно. В рамках схемно-целевого подхода выделяют две схемы: детерминистскую (обусловленную) и стохастическую (вероятностную). Схема определяется в зависимости от типа исследуемых данных и определяет используемые в исследовании методы. Наиболее трудным остается вопрос выбора соответствующих условий проведения эксперимента. Например, при анализе совместной встречаемости особей двух видов растений (животных) необходимо выбрать элементарный контур (круг, квадрат или прямоугольник), который «случайно бросается» на обследуемый участок. При анализе зависимости категорий растительности от условий среды также возникает проблема ранжирования факторов среды на ряд категорий. После получения выборочных конечных схем производятся выбор и расчет статистических мер, а также и оценка ошибок выборочности. Но и в этой области было разработано достаточно большое число методов. Далее производим сбор данных. Для того чтобы осуществить эту процедуру, прежде всего, необходимо осуществить выбор элементарного описания. Выбор элементарного описания производится на основе принципа «воспроизводимости», а также принципа «разделимости» описаний. Суть вышеуказанных принципов состоит в следующем: «Первый принцип предполагает воспроизводимость описаний другими исследователями. Для этого следует чётко определить наименьшее количество признаков и их категорий, по которым производится описание объектов. Эти признаки и их категории должны быть доступны для восприятия многими исследователями. Затем выбирается «элементарное описание», исходя из второго принципа – разделимости. По этому принципу, элементарные описания одного и того же объекта должны быть сходными, а описания существенно различных объектов – различными» [47] Проблема выбора элементарного описания неоднократно рассматривалась в работах ведущих специалистов. Основными вопросами были обсуждение вопросов формы и размеров элементарного описания (они должны быть приблизительно равны и ограничены, причём последнее условие зависит от типа экосистемы и её географической составляющей). Ежё одно условие, указанное А.И. Толмачевым, требует высокой степени изученности сравниваемых флор. Все эти вопросы подробно изложены в литературе по экологии и смежным наукам. Основным уровнем изучения таксономического разнообразия является видовой. Совокупность видов любого биоценоза формально можно представить конечным множеством, булевым вектором (набором) или точкой многомерного пространства Хэмминга (см. например, [48]). Флористические данные обычно представляются в форме прямоугольной таблицы размером n Понятие «биологическое разнообразие» за весьма короткий отрезок времени получило большое количество толкований. В биологическом смысле рассматриваются представления о внутривидовом, видовом и надвидовом (ценотическом) разнообразии жизни. Однако, в добавление к этому, сначала деятели охраны природы, а затем и ученые стали говорить об экосистемном и ландшафтном разнообразии как объектах сохранения, а, соответственно, изучения и выделения в природе. Особенно отметим работы Р. Уиттекера, в которых была предложена организация уровней экосистемного разнообразия и исследованы зависимости биоразнообразия от факторов окружающей среды. Схематически совокупность уровней биоразнообразия F(R) можно представить в виде:
где нижний ряд греческих букв обозначает инвентаризационное разнообразие (внутри объекта). Например для растительного покрова: w(омега) – разнообразие планетарной флоры; c(хи) – разнообразие флористического царства; u(ипсилон) – разнообразие флористической области; s(сигма) – разнообразие флористической провинции; p(пи) – разнообразие флористического округа; x(кси) – разнообразие флористического района; m(мю) – разнообразие конкретной флоры. Верхний ряд букв соответствует дифференцирующие разнообразию. Для растительного покрова соответственно: y(пси) – разнообразие между флористическими царствами в пределах флоры планеты; f(фи) – разнообразие между флористическими областями в пределах царства; t(тау) – разнообразие между флористическими провинциями в приделах флористической области; r(ро) – разнообразие между флористическими округами в пределах флористической провинции; o(омикрон) – разнообразие между флорами районов в пределах флористического округа; n(ню) – разнообразие между конкретными флорами в пределах флористического района. Для оценки инвентаризационного разнообразия используются такие меры как мера Шеннона, индекс Маргалефа, индекс Бриллюэна, индекс Симпсона и др. Рассмотрению особенностей этих мер посвящено немало публикаций [32, 40, 55 и др.]. Приведём меру К. Шеннона:
где pi – это доли обилия каждого вида в сообществе. Теоретически Н -функция принимает максимальное значение тогда, когда имеет место полная выравненность распределения, что соответствует наибольшей сложности систематической структуры. Подход к флорам и фаунам как дескриптивным множествам видов или их групп (крупных таксонов, элементов флоры) позволяет использовать для сравнительного изучения флористических систем весьма простой, но вполне строгий математический аппарат алгебры дескриптивных множеств, описанный нами выше. Он отражает (при сравнении различных структур флоры) пропорциональный вклад каждого таксона и элемента, а не только его порядковый номер в нисходящем ряду. Напротив, коэффициенты ранговой корреляции, часто используемые при сравнении таксономических и иных структур флоры, при большей трудоемкости их расчета огрубляют выражение соотношений таксонов или элементов, переводя его из шкалы отношений в более слабую порядковую шкалу. В 1973 году Б.И. Семкин предложил абсолютную меру разнообразия, основанную на сравнении исследуемого весового множества с эталоном, имеющим максимальное разнообразие:
где Прежде чем перейти к блоку анализа данных в рамках схемно-целевого подхода, необходимо чётко представлять себе суть используемых аналитических методов. Используя сравнительный анализ объектов, мы определяем отношения «целого-части», которые могут быть равноправными (определяются симметричные отношения конвергенции или дивергенции), неравноправные (определяются отношения направленной конвергенции или дивергенции) и различные комбинации отношений, требующие определения симметричных и не симметричных отношений конвергенции или дивергенции. Сёмкиным Б.И. в ряде работ были разработаны теоретические основания введения мер близости, используемых в экологии и других смежных областях. Важным следствием из данной теории является понятие эквивалентности мер (которое позволило объединить множество разных мер в одну теорию). Например, общая формула для мер сходства (в общем виде – меры конвергенции) выглядит следующим образом:
где Вернёмся к схеме анализа. Ha основе матрицы данных легко можно рассчитать матрицу мер конвергенции, что, например, для детерминистской модели соответствует матрице абсолютных мер сходства (или матрице пересечения) размером q2:
т.е. для каждой пары объектов определяется пересечение, которое соответствует количеству общих (для обоих объектов) видов. Под диагонали матрицы располагается общее число видов одного объекта (или на обилие). Далее определяем степень расхождения объектов по числу видов: если объекты разновелики – применяем в основном меры включения, если равновелики – меры сходства (или различия). Также в некоторых случаях возможно применение смешанного анализа. Для ненормированных данных матрица пересечения легко преобразуется в матрицу мер включения. Приведём одну из возможных мер включения:
где K0 (a; b) – мера включения b в a; K0 (b; a) – мера включения a в b. Т.е. меры включения указывают на степень вхождения одного объекта в другой по видовому составу. В целом стоит отметить, что меры включения более информативны в сравнении с мерами сходства, т.к. последние являются осреднением мер включения. Получить матрицу включения из матрицы пересечения очень просто. Для этого необходимо элементы каждой строки матрицы пересечения разделить на соответствующий этой строке диагональный элемент. В полученной матрице включения целесообразно указывать направление включение, т.к. матрица включения несимметрична. Визуализировать отношения включения можно с помощью ориентированных графов. Для их построения требуется задать некоторый порог включения. Далее, просматривая строки матрицы включения, отмечаем все числа, которые превышают пороговую величину. Вершину графа с номером строки соединяем с вершиной графа номера столбца, на пересечении которого и стоит отмеченное число. Вершинами графа будут названия районов. Граф включения показывает связь флористических участков (по сходству видов). Матрицу включения можно привести к симметричному виду, рассчитав меру сходства для каждой пары объектов, или используя меру сходства (например, меру Сёренсена):
где Kαβ и Kβα – несимметричные меры включения. Отношения сходства обычно визуализируются с помощью графического представления метода иерархического кластерного анализа – дендрограммы. Помимо выбора меры сходства или меры различия для матрицы необходимо также выбрать способ построения дендрограммы. Кроме перечисленных алгоритмов упорядочивания данных также применяются методы построения диаграммы Чекановского, дендрита (неориентированный граф) и др. После анализа следует этап интерпретации. Исследователь, исходя из системного понимания территории и взаимосвязей внутри объекта, адекватно интерпретирует полученные результаты. Из-за высокой сложности исследуемых объектов к процессу исследования, как правило, подключаются специалисты разных направлений – ботаники, зоологи, биогеографы, геоботаники, фаунисты, экологи, математики, программисты и др.
Вопросы для самоконтроля 1. Назовите объекты биологического мониторинга. 2. Биоиндикация как метод оценки загрязнения окружающей среды. Приведите примеры. 3. Оценка биологического разнообразия. Что такое биоразнообразие? Основные показатели таксономического разнообразия и их информативность. 4. Количественная оценка биологических объектов. Опишите концепцию основных уровней биоразнообразия по Уиттеккеру. Расскажите об основных индексах оценки инвентаризационного и дифференцирующего разнообразия.
|
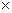 m, где n – количество объектов (например, флор), a m – количество признаков (например, видов) в списке. Такая таблица обозначается различными терминами в литературе: матрица наблюдений, матрица флористических (или фаунистических) данных, «первичная матрица» (primary matrix) или в определённых условиях – «сводный список». Форма представления чисел в матрице данных даёт нам первичную информацию об объекте и дальнейших возможностях использования математических методов. Так, например, булевы наборы дают нам информацию только о присутствии-отсутствии вида во флоре, тогда как дескриптивные множества дают некоторое представление об обилии видов. Мы также можем сравнивать биологические спектры (например спектры Раункиера) и другие разновидности данных, например, такие как случайные события или случайные величины. В современной науке есть несколько алгоритмов обработки первичной матрицы. Анализ матрицы n
m, где n – количество объектов (например, флор), a m – количество признаков (например, видов) в списке. Такая таблица обозначается различными терминами в литературе: матрица наблюдений, матрица флористических (или фаунистических) данных, «первичная матрица» (primary matrix) или в определённых условиях – «сводный список». Форма представления чисел в матрице данных даёт нам первичную информацию об объекте и дальнейших возможностях использования математических методов. Так, например, булевы наборы дают нам информацию только о присутствии-отсутствии вида во флоре, тогда как дескриптивные множества дают некоторое представление об обилии видов. Мы также можем сравнивать биологические спектры (например спектры Раункиера) и другие разновидности данных, например, такие как случайные события или случайные величины. В современной науке есть несколько алгоритмов обработки первичной матрицы. Анализ матрицы n 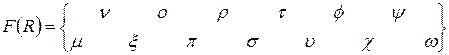 ,
, ,
, ,
, ; X – весовое множество, разнообразие которого определяется; n – число таксонов. Некоторые аналогичные идеи были изложены в работах других исследователей. Большинство из них сравнивали определённые объекты с идеальным объектом, но также и существует идея использования некоторых естественных эталонов. Одним из таких эталонов может являться климаксное сообщество и, следовательно, упомянутые меры можно использовать для оценки сукцессионной динамики, а также для определения уровня антропогенной нагрузки на экосистему.
; X – весовое множество, разнообразие которого определяется; n – число таксонов. Некоторые аналогичные идеи были изложены в работах других исследователей. Большинство из них сравнивали определённые объекты с идеальным объектом, но также и существует идея использования некоторых естественных эталонов. Одним из таких эталонов может являться климаксное сообщество и, следовательно, упомянутые меры можно использовать для оценки сукцессионной динамики, а также для определения уровня антропогенной нагрузки на экосистему.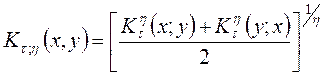 ,
,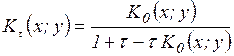 ,
, 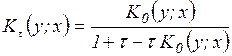 ,
, 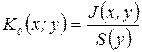 ,
, 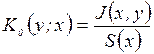 ,
, 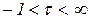 ,
,  . При
. При 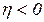 ,
, 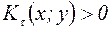 и
и 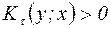 . Все меры, упорядоченные параметром τ; являются эквивалентными, а упорядоченные параметром η; – неэквивалентными. При η=-1 получаем класс средней гармонической средней, в который входят известные меры Сёренсена (τ=0), Жаккара (τ=1). Меры сходства двойственны мерам различия (известным как расстояния), т.е. эти меры взаимно дополняют друг друга (если объекты на 20% сходны, то очевидно, что они на 80% различны).
. Все меры, упорядоченные параметром τ; являются эквивалентными, а упорядоченные параметром η; – неэквивалентными. При η=-1 получаем класс средней гармонической средней, в который входят известные меры Сёренсена (τ=0), Жаккара (τ=1). Меры сходства двойственны мерам различия (известным как расстояния), т.е. эти меры взаимно дополняют друг друга (если объекты на 20% сходны, то очевидно, что они на 80% различны).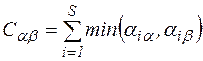 ,
, ;
;  ,
,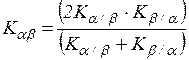 ,
,


