Теория боли
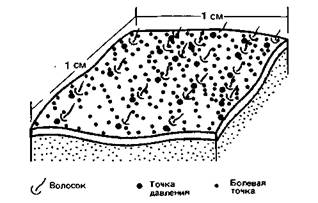
Теория специфичности боля. Современные гипотезы о происхождении боли в тканях исходят из того, что это независимое ощущение со своим собственным специализированным нервным аппаратом из рецепторов, проводящих путей и центров. Согласно такому представлению, подкрепленному многими экспериментальными данными, все люди и практически все животные обладают особыми рецепторами с очень высоким порогом, которые возбуждаются только стимулами, повреждающими или грозящими повредить окружающую ткань. Рецепторы, реагирующие на такие «вредные» стимулы, названы ноцицепторами,а активируемые ими нейронные структуры - ноцицептивной системой. Соответственно рецепция, проведение и центральная нервная обработка вредоносных сигналов составляют иоцицепцию;тем самым проводится грань между «объективными» нейронными процессами и «субъективным» ощущением боли. С такой точки зрения «теория специфичности боли» становится просто теорией специфичности ноцицепции. Это лишь один пример иногда еще встречающегося смешения терминов «ноцицепция» и «боль». Однако их следует различать: ноцицептивные нейронные структуры и происходящие в них электрические и химические процессы отнюдь не эквивалентны субъективно испытываемой боли. Одним из первых экспериментальных подтверждений теории специфичности было наблюдение, проиллюстрированное рис. 10.4: болевая чувствительность не распределена по коже равномерно; как и в случае механо- и терморецепции, болевые стимулы воспринимаются только в дискретных болевых точках. Их гораздо больше, чем точек давления (отношение 9:1 на рис. 9.4). Холодовых и тепловых точек на коже еще меньше, чем последних. Уже по этой причине представлялось вероятным, что ноцицепция обслуживается особыми, специализированными ноцицепторами, а не механо- и терморецепторами, как требовали бы описанные ниже теории интенсивности и распределения импульсов. Ноцнцептивные теории интевсивиости и распределения импульсов (pattern theory). Долгое время эти теории конкурировали с теорией специфичности. Они исходили из того, что огромное разнообразие вредных стимулов (т. е. отсутствие единственного адекватного стимула) как будто несовместимо с существованием специализированных ноцицепторов. Согласно этим концепциям, боль возникает
всегда, когда интенсивность стимуляции низкопороговых механо- и терморецепторов превышает определенный уровень. По теории интенсивности ноцицептивные стимулы вызывают особо высокочастотные вспышки импульсацни низкопороговых рецепторов, а теория распределения импульсов предполагала особый характер импульсацни,отличный от возникающего в ответ на безвредные стимулы. Эти теории привлекали главным образом своей кажущейся простотой, поскольку связывали ноцицепцию со специфическими нейронными структурами. В наше время они уже не представляются такими простыми, как прежде; кодирование разной сенсорной информации в одной нервной сети считается сейчас значительно более сложной проблемой, чем ее обработка в разных системах. Теория воротного контроля. Предложенная в 1965 г. Мелзаком и Уоллом [22] теория воротного контроля спинальной переработки ноцицептивной информации постулировала, что торможение относящихся к ноцицептивной системе центростремительных нейронов задних рогов обусловлена возбуждением толстых неноцицептивных афферентов (ворота закрыты), а активацию ихвозбуждением тонких ноцицетпивных афферентов (ворота открыты). Считалось, что такое торможение генерируется в студенистом веществе заднего рога спинного мозга и (это было главным в теории) обеспечивается исключительно пресинаптическbм тормозным механизмом,действующим на топкие ноцицептивные афференты. Данную гипотезу не удалось подтвердить экспериментально, ее основные положения были отвергнуты [26, 27], и авторам пришлось внести в свою концепцию изменения [8]. Второй основной постулат теории воротного контроля состоит в том, что спинальные тормозные механизмы ноцицепции в студенистом веществе активируются также нисходящими тормозными системами,т. е. даже на спиналъном уровне ноцицептивная информация находится под центробежным контролем. Наличие таких нисходящих тормозных систем теперь считается доказанным не только в ноцицеп- ГЛАВА 10 НОЦИЦЕПЦИЯ И БОЛЬ 227 тивной, но и во всех других соматосенсорных системах, причем в некоторых из них они известны уже давно (см. рис. 9.26, с. 219). Заслуга теории воротного контроля остается в том, что она привлекла внимание к существенной модуляции ноцщептивных входов в спинной мозг на уровне уже первых центральных нейронов локальными и нисходящими влияниями [14,27, 30].
|